|
|
| Le financement de ce site est assuré par vos dons, merci! | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|

| |
Après « L’erreur de Descartes », voici « L’erreur de Broca » Une première étude d’imagerie cérébrale sur les effets du LSD L’étrange ordre des choses selon Antonio Damasio « Moi » conscient versus motivations inconscientes
Cognition et émotions incarnées Nos sentiments pourraient se passer de l’insula La contribution du claustrum au sentiment d’être soi
| |
Des modèles neurobiologiques de la conscience, comme celui de l’espace de travail global, prédisent que les contenus conscients sont largement distribués dans le cerveau. C’est ce que confirment de nombreuses expériences en imagerie cérébrale, notamment celles de Stanislas Dehaene et ses collaborateurs, qui ont observé que l’allongement de la durée de présentation d’un mot sur un écran juste au-delà de son seuil de perception conscient s’accompagne d’une augmentation importante de l’activité des aires frontales, préfrontales, cingulaires antérieures et pariétales. Les inputs sensoriels conscients produiraient donc une activité cérébrale bien plus étendue que des stimuli inconscients comparables et l’activation soudaine des lobes frontaux et pariétaux serait la signature typique d’une perception consciente. Mais cette conscience perceptuelle, certains diraient cette conscience primaire, n’est pas le seul sens que l’on donne au mot conscience. Il est donc nécessaire de bien définir de quel niveau de conscience on parle quand on essaie de lui associer des structures cérébrales. Par exemple, la première condition nécessaire pour qu’il y ait possibilité de traitement conscient des stimuli sensoriels extérieurs est d’être dans un état de vigilance approprié (par exemple être éveillé plutôt qu’endormi).
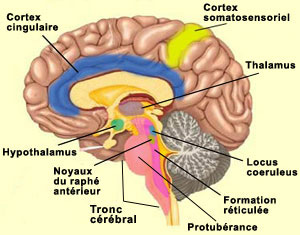
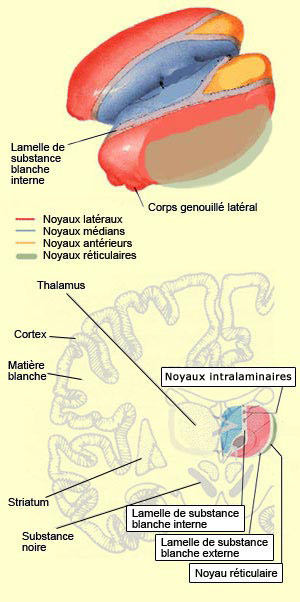
Il faut noter ici que l’activité de la formation réticulée, tout comme celle des aires sensorielles primaires, semble être nécessaire mais non suffisante pour une expérience consciente plus élaborée. Cet autre niveau est atteint avec ce que plusieurs appellent la conscience primaire, c’est-à-dire un état de veille où nous sommes en relation avec notre environnement « ici et maintenant ». À la suite des travaux de Bjorn Merker, il semble que le tronc cérébral joue un rôle plus important qu’on ne l’aurait cru dans cette conscience primaire. Damasio désigne pour sa part ce niveau de conscience sous l’expression de « conscience noyau ». Celle-ci dépendrait selon lui principalement du cortex cingulaire et des noyaux intralaminaires du thalamus. Des expériences ont en effet démontré que la destruction bilatérale de la partie centromédiane des noyaux intralaminaires du thalamus abolit la conscience, produit un coma, ou cause d’autres états proches de la mort cérébrale. De plus, cette région du thalamus est également l’un des principaux sites d’action des substances anesthésiantes et des drogues anti-psychotiques. Ce n’est pas d’hier que le thalamus est impliqué dans des modèles de la conscience. Déjà dans l’une des premières hypothèses sur la conscience, celle de Francis Crick en 1984 et de son hypothèse du projecteur thalamique (“thalamic searchlight hypothesis”, en anglais), le thalamus contrôlait quelle région du cortex devenait le point focal de la conscience. Une idée semblable, mais plus sophistiquée, était reprise plus récemment par Rodolfo Llinas qui fait l’hypothèse que les oscillations de certains neurones thalamiques servent en quelque sorte de rythme de base sur lequel se synchronise les oscillations corticales des différentes modalités sensorielles pour former une image unifiée de l’environnement. Un peu comme un chef d’orchestre qui donne la mesure à ses musiciens (voir l'encadré). Une solution originale au problème de liaison ou, selon l’expression anglaise consacrée, du « binding problem ».
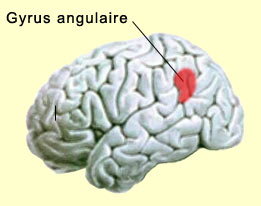
Ces « boucles thalamo-corticales » en sont venues à jouer un rôle important dans pratiquement toutes les théories neurobiologiques qui tentent d’expliquer les états de conscience supérieurs. Car les niveaux de conscience inférieurs dont on vient de parler ne sont en quelque sorte que des conditions préalables aux niveaux supérieurs de la conscience humaine, ceux de la conscience réflexive et de la conscience de soi. La conscience réflexive, cette impression que « c’est moi qui perçois », est souvent présentée comme condition nécessaire à la conscience de soi, c’est-à-dire le sentiment d’être soi-même et pas un autre. Cette dimension autobiographique implique que nous puissions nous représenter des expériences conscientes dans le passé ou le futur, et nécessite donc la contribution de la mémoire et de nos fonctions supérieures permettant la conceptualisation abstraite et la planification. On peut donc s’attendre à ce que les régions cérébrales reconnues pour être impliquées dans ces fonctions, en particulier dans les lobes frontaux et pariétaux, soient sollicitées par cette conscience de soi. Et c’est ce que montrent effectivement certaines études qui se sont penchées spécifiquement sur cette question. Ceci dit, il semble que ces niveaux supérieurs de la conscience impliquent également d’autres structures cérébrales dont le rôle demeura longtemps méconnu, entre autres parce que certaines d’entre elles sont situées en profondeur dans le cerveau, ce qui les rendaient difficilement accessibles. Une difficulté qui a pu être contournée avec les techniques d’imagerie cérébrale.
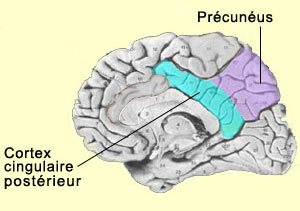
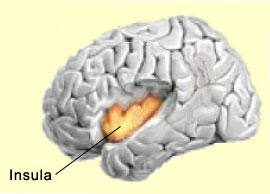
Cette hypothèse s’accorde aussi avec les études ayant démontrée une baisse d’activité dans le cortex pariétal postéro-médian dans de nombreux états de conscience altérée, comme le sommeil, l’anesthésie ou l’état végétatif. D’autres études ont également mis en évidence une baisse d’activité dans le précunéus et le cortex cingulaire postérieur lorsque le sujet est sous hypnose, un état de conscience également modifiée. Enfin, le précunéus semble aussi jouer un rôle dans l’imagerie visuo-spatiale. Certaines expériences montrent par exemple que le précunéus est plus actif durant l’imagerie motrice d’un mouvement du doigt que durant la réelle exécution de ce mouvement, ce qui semble indiquer encore ici une propension à se représenter son propre corps dans l’espace. L’insula est une autre région du cerveau longtemps restée méconnue à cause de sa position en profondeur dans les replis du cortex. Et comme elle n’était pas non plus associée aux fonctions dites « supérieures », cela la rendait moins intéressante aux yeux de ceux qui s’intéressaient à l’étude de la conscience. Mais cette indifférence s’est transformée en un vif regain d’intérêt à la suite des travaux de Antonio Damasio et de son hypothèse des marqueurs somatiques qui fait la part belle à l’insula. Damasio a en effet proposé que cette région du cortex permettrait de cartographier nos états viscéraux qui sont associés à des expériences émotionnelles, donnant ainsi naissance à un sentiment conscient. Ces travaux s’inscrivent dans le courant de la cognition incarnée où la pensée rationnelle consciente ne peut être séparée des émotions et de leur incarnation dans le reste du corps.
L’insula est aussi déjà bien associée aux processus de douleur ainsi qu’à plusieurs émotions de base comme la colère, la peur, le dégoût, la joie ou la tristesse. Sa portion la plus antérieure est considérée comme faisant partie du système limbique. L’insula serait aussi grandement impliquée dans les désirs conscients, comme la recherche active de nourriture ou de drogue. Ce qu’il y a de commun dans tous ces états, c’est qu’ils affectent le corps entier en profondeur. Un constat qui tend à renforcer son rôle probable dans la représentation que nous nous faisons de notre propre corps ainsi que dans l’aspect subjectif de l’expérience émotionnelle. Enfin, l'insula humaine, et à un moindre degré celle des grands singes, aurait deux innovations évolutives qui lui permettrait de porter la lecture de notre état corporel à un niveau inégalé chez les autres mammifères. D'abord la partie antérieure de l'insula, et plus particulièrement de l'insula de l'hémisphère droit, serait davantage développée chez les humains et les grands singes que chez les autres espèces animales. Ceci permettrait un décodage plus précis de nos états viscéraux, et donc par exemple à une simple mauvaise odeur de devenir un sentiment de dégoût, ou encore au toucher d'une personne aimée de se transformer en sentiment de délice. L'autre
modification majeure à notre insula est la présence d'un type de
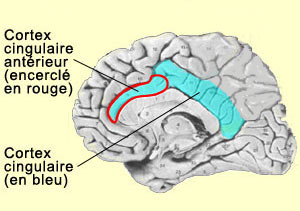
neurone que l'on retrouve seulement chez les grands singes et l'humain. Il s’agit de grandes cellules nerveuses allongées en forme de cigare appelées VEN. De plus, on ne retrouve ce type de neurone que dans l’insula et le cortex cingulaire antérieur. Ces neurones font des connexions avec diverses parties du cerveau, ce qui serait un atout essentiel pour les fonctions supérieures qu’on attribue à ces deux structures cérébrales. Lorsqu’on pique un sujet avec une aiguille, son cortex cingulaire augmente son activité, si bien qu’on réfère souvent à ces neurones comme aux « neurones de la douleur ». Détail pour le moins intéressant, William Hutchison et ses collègues de l'Université de Toronto ont montré en 1999 que ces mêmes neurones du cortex cingulaire vont également s’activer lorsque le sujet voit quelqu’un se faire piquer avec une aiguille. Pour ce type de neurones, appelés neurones miroirs, il n’y a donc pas de frontière entre soi et l’autre… Les primates, et donc les humains, sont des créatures hautement sociales. Connaître les intentions des autres a de tout temps été crucial pour notre survie. Voilà pourquoi nous sommes passé maître, peut-être avec l’aide des neurones miroirs, dans l’art de la simulation interne de l’esprit des autres. Certains, comme V.S. Ramachandran, suggèrent même que cette capacité à décoder les états d’esprits d’autrui aurait été la première à évoluer et qu’elle aurait été par la suite appliquée à nous-même, pour devenir ce que nous appelons la conscience de soi. Et pour Ramachandran, non seulement les neurones miroirs mais toutes les aires qui contribuent au langage, comme celle de Wernicke située dans le lobe temporal par exemple, seraient des joueurs incontournables dans cette aventure. Une importance du langage que l’on retrouve dans plusieurs modèles de cette conscience supérieure, dont celui de Michael Gazzaniga, connu pour son travail avec les patients au «cerveau divisé». Mais si Gazzaniga localise dans l’hémisphère du langage «l’interprète» qui nous rendrait conscient de nous-même, d’autres comme Edelman affirment que la conscience n’est le fait d’aucune structure précise du cerveau. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

| |
|
|
|
|
|
|
|
|