L’ophtalmoscope,
l’appareil servant à regarder le fond de l’oeil à travers
la pupille, permet de voir les vaisseaux sanguins à la surface de la rétine.
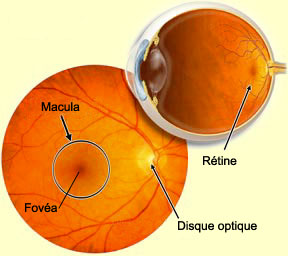
Ces vaisseaux sanguins pénètrent dans l’œil en passant
par une région circulaire décolorée de la rétine appelée
le disque optique. C’est aussi à cet endroit, qui
constitue la tête du nerf optique, que les axones des cellules
ganglionnaires se rejoignent pour sortir du globe oculaire.
Étant
donné l’absence de photorécepteurs
à cet endroit, l’œil y est insensible à la lumière,
tout comme d’ailleurs à l’endroit où passent les plus
gros vaisseaux sanguins. C’est pour cette raison que le disque optique reçoit
aussi l’appellation de point aveugle. Pourtant, nous ne
ressentons pas d’interruption dans notre champ visuel au point aveugle car
le cerveau « compense » d’une certaine façon la
perception visuelle à ces endroits (voir la capsule expérience
à gauche).
Au centre de la rétine se trouve une
partie plus sombre, la macula, qui est pratiquement
dépourvue de vaisseaux sanguins afin d’optimiser la
vision centrale (par opposition à la vision périphérique).
Au centre de la macula, une petite dépression d’environ
2 mm de diamètre forme la fovéa,
le point de la rétine constitué exclusivement de cônes
où l’acuité visuelle est la meilleure.
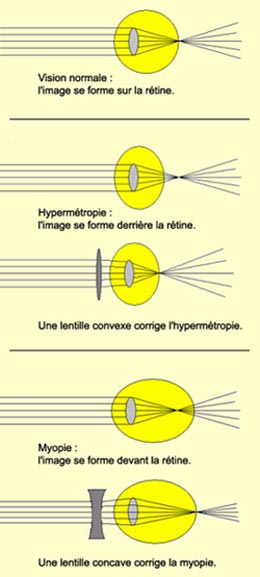
Plusieurs troubles
de la vision peuvent surgir quand les rayons lumineux ne convergent pas exactement
sur la rétine.
Si le globe oculaire est trop court d’avant
en arrière par exemple, les rayons convergent au-delà de la rétine.
Ce défaut appelé hypermétropie se corrige
en plaçant une lentille convexe devant l’œil, lentille qui va
accentuer la convergence et ramener le foyer sur la rétine.
Dans
le cas de la myopie, la convergence se fait en avant de la rétine
lorsque le globe oculaire est trop long par exemple. On corrige la myopie par
le port de verres concaves.
Un autre trouble de la vision, la
presbytie, vient d’un durcissement du cristallin lié
à l’âge. Ce durcissement diminue l’élasticité
du cristallin, l’empêchant de prendre une forme suffisamment arrondie
lors de l’accommodation,
et de s’aplatir assez au cours de la relaxation. La correction de la presbytie
fait appel à une lentille bifocale, la partie supérieure étant
concave pour la vision de loin, et la partie inférieure convexe pour la
vision de près.
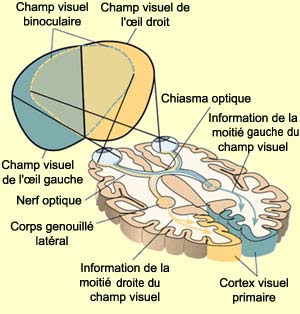
Le champ visuel d’un œil
est la région de l’espace qui vient s’imprimer sur la rétine
de cet œil lorsque le regard est fixé sur un point éloigné.
Les champs récepteurs des deux yeux se recoupent en grande partie, mais
celui de l’œil droit est plus large à droite, et celui de l’œil
gauche à gauche.
La figure ci-dessus montre aussi comment la partie droite
du champ visuel (en jaune) se retrouve analysée dans
le cortex
visuel gauche, et vice versa. La partie centrale de l’espace
couverte par les deux yeux constitue le champ visuel binoculaire.
Les nombreuses connexions qui relient les différentes
régions cérébrales impliquées dans le traitement de
l’information visuelle (cortex visuel, ganglions
de la base, protubérance, cervelet,
noyaux occulomoteurs, etc.) sont généralement réciproques.
Cette réciprocité crée des boucles de rétroaction
qui montrent bien à quel point le système visuel est intrinsèquement
lié au fonctionnement de l’ensemble du système
nerveux.
La présence de ces nombreuses boucles de rétroaction
neuronales dans le système visuel prend tout son sens à la lumière
de l’évolution.
Dans les cerveaux
reptiliens par exemple, la vision est très liée à des
réponses réflexes de défense de l’organisme. Un bruit
assourdissant, une nouvelle sensation tactile ou un objet qui approche rapidement
de nous par le côté amène l’organisme à tourner
la tête rapidement vers le nouveau stimulus pour que les yeux puissent en
évaluer le danger réel. Même si le système visuel des
primates s’est beaucoup perfectionné pour nous permettre une vision
consciente et détaillée du monde qui nous entoure, ces vieux circuits,
toujours utiles, ont été conservés par l’évolution
et sont encore fonctionnels dans le cerveau humain.
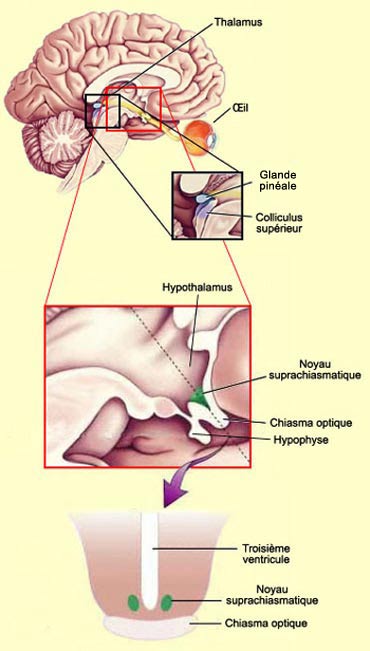
C’est le cas de
l’hypothalamus, et plus précisément de son
noyau suprachiasmatique, qui reçoit un certain nombre
de connexions des axones en provenance de la rétine. Le noyau suprachiasmatique
est considéré comme le site majeur de notre horloge biologique interne.
C’est pourquoi être informé en permanence de la clarté
ou de l’obscurité ambiante lui permet de synchroniser toute une série
de rythmes biologiques liés au cycle quotidien du jour et de la nuit, y
compris le sommeil
et l’éveil.
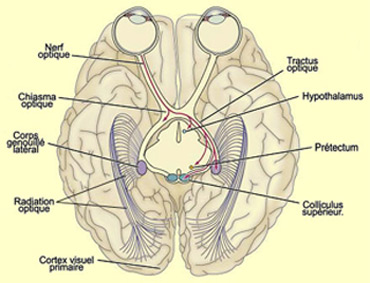
D’autres axones des cellules ganglionnaires
de la rétine vont également rejoindre le prétectum,
une partie du mésencéphale qui contrôle l’ouverture
de la pupille et certains mouvements des yeux.
Enfin, environ 10%
des axones rétinofuges se projettent sur une partie du tectum (ou toit)
du mésencéphale appelée colliculus supérieur.
Cette voie est relativement importante, considérant qu’elle comporte
tout de même environ 150 000 axones, l’équivalent du nombre
total de cellules ganglionnaires rétiniennes chez le chat ! En fait, le
colliculus supérieur correspond au tectum optique de tous les vertébrés
non mammifères chez qui cette projection rétinotectale est la voie
rétinofuge principale.
À cause de la superposition des
champs
récepteurs dans la rétine, la projection d’un point lumineux
sur celle-ci active une grande population de neurones du colliculus supérieur.
Ces derniers provoquent des mouvements des yeux et de la tête, par l’intermédiaire
de neurones
moteurs du tronc cérébral, qui vont chercher à amener
l’image du spot lumineux sur la fovéa.
La voie rétinotectale est donc impliquée dans l’orientation
du regard quand l’œil est sollicité par un stimulus dans la
périphérie du champ visuel.
À l’instar du
corps genouillé latéral, le colliculus supérieur reçoit
également des connexions du cortex visuel primaire. Les cibles des neurones
du colliculus supérieur sont pour leur part des structures sous-corticales,
comme la formation réticulée, le colliculus inférieur et
la moelle épinière. Les neurones du colliculus supérieur
ont aussi une influence sur deux structures qui sont impliquées dans la
vision : le corps genouillé latéral et le pulvinar.
Le pulvinar
est un noyau de la région postérieure du thalamus qui reçoit
des afférences d’axones collatérales directement du tractus
optique ainsi que par l’entremise du CGL. Comme plusieurs noyaux thalamiques,
il a longtemps été considéré comme un relais plutôt
passif de l’information en route vers le cortex où le véritable
traitement de l’information aurait lieu. Mais l’accumulation de données
montrant que les neurones du pulvinar ont des réponses visuelles sophistiquées
que l’on croyait réservées au cortex, a radicalement changé
cette conception.
Le pulvinar serait plutôt un centre d'interprétation
de l'image qui jouerait un rôle important dans l'attention visuelle et dans
la perception du mouvement. Il contribuerait par exemple au maintien de la stabilité
de notre environnement visuel en compensant l’effet qu’ont les mouvements
du corps sur le positionnement des images sur la rétine. Il n’est
donc pas surprenant qu’on ait découvert que les neurones du pulvinar
projettent vers les aires visuelles secondaires
impliquées dans la détection du mouvement.
L'agnosie
visuelle, qui survient souvent à la suite d'une lésion
occipito-temporale bilatérale, rend le sujet incapable de discriminer la
forme des objets, même s’il conserve parfois de bonnes capacités
de discrimination des couleurs et des textures. Mais il reconnaît très
mal la forme d'objets géométriques arbitraires, les lettres de l'alphabet
et des dessins en noir et blanc. En revanche, il peut très bien diriger
efficacement sa main vers un objet, orienter son poignet pour introduire un objet
dans une fente et disposer ses doigts de manière à saisir un objet.
Le syndrome d’agnosie visuelle est donc un déficit de la conscience
visuelle mais où l'aptitude à manipuler des objets est conservée.
À l’opposé, dansl'ataxie optique, des patients souffrant de lésions
au lobe pariétal sont capables de reconnaître les objets mais pas
de les prendre et de les utiliser convenablement. Ce genre d’observation
a permis de mettre en évidence les rôles différents des voies
visuelles ventrales et dorsales.
En 1983, Joseph Zihl et ses collaborateurs ont
publié à Munich un article consacré à une femme de
43 ans qui était devenue totalement incapable de percevoir les mouvements
à la suite d’un accident vasculaire cérébral qui avait
lésé les deux côtés de son cortex extrastrié
impliqué dans la reconnaissance du mouvement (aire V5). Cette patiente
souffrait donc de l’étrange syndrome de la cécité
au mouvement (ou akinétopsie) qui se manifeste par des "arrêts
sur image" de plusieurs secondes tout au long desquels elle ne perçoit
qu’une image immobile en perdant toute conscience visuelle des mouvements
dans son environnement. Traverser une rue était par exemple fort périlleux
pour cette patiente puisqu’une voiture qu’elle avait vue "arrêtée"
à une grande distance de l’endroit où elle était pouvait
se retrouver tout près d’elle après qu’elle eut commencé
à traverser. Se verser un verre d’eau pouvait être tout aussi
problématique puisqu'elle voyait l’eau qui coule comme gelée
et qu’elle comprenait qu’elle en avait trop versée quand elle
découvrait soudain l’eau répandue sur la table.
LES DIFFÉRENTS CORTEX VISUELS
Suite aux travaux fondateurs
de Leslie Ungerleider et Mortimer Mishkin publiés en 1982, on distingue
deux grandes voies dans le traitement cortical de l’information visuelle
: la voie ventrale, celle de l’identification de l’objet (le «
quoi ? »), et la voie dorsale, celle de la position de l’objet (le
« où ? »). Divers travaux ultérieurs ont toutefois amené
quelques remises en question de cette dichotomie. C’est le cas, entre autres,
des études où chacune des voies était sélectivement
détériorée chez le singe ou encore celles avec des humains
ayant subi des lésions cérébrales n’ayant touché
qu’une seule des deux voies (voir encadrés).
On pense aujourd’hui
que la fonction principale de la voie dorsale est de guider en
temps réel les actions que nous dirigeons vers des objets du monde visuel.
De plus, le traitement réalisé par cette voie serait largement inconscient.
La voie dorsale serait une « voie de l’action » car en intégrant
les relations spatiales entre l’organisme et l’environnement, elle
nous permet d’interagir efficacement avec lui.
La
voie ventrale semble quant à elle plutôt impliquée dans
le formation de représentations conscientes de l’identité
des objets. À la dichotomie fonctionnelle entre voie dorsale et ventrale
se doublerait donc une autre opposition, celle de la vision consciente versus
la vision inconsciente.
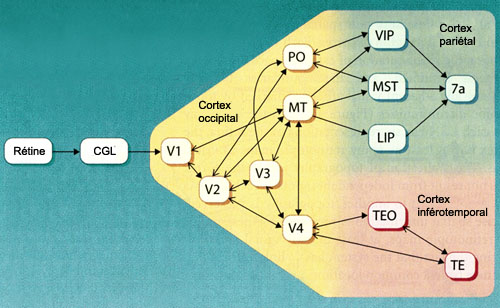
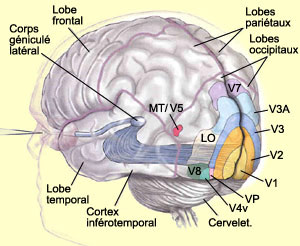
La
voie dorsale comprend plusieurs régions corticales dont l’aire temporale
médiane (MT ou V5), l’aire temporale médiane supérieure
(MST) et les aires intrapariétales ventrales et latérales (VIP et
LIP).
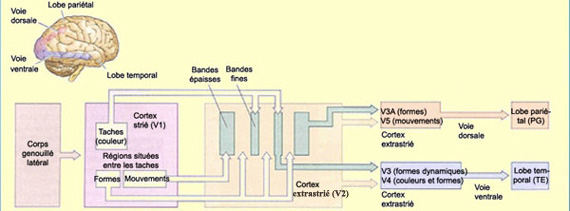
L’aire V5 (aussi appelée aire MT)
semble contribuer de façon importante à la perception du mouvement.
Cette région reçoit des projections des aires V3, V2 ainsi que de
la
couche IV-B de l’aire visuelle primaire (V1). Il est intéressant
de noter que cette couche IV-B fait justement partie du canal magnocellulaire
impliqué dans l’analyse du déplacement des objets. Ce canal
conserve d’ailleurs sa spécificité pour le mouvement dans
l’aire V2 où il se concentre dans les stries larges riches en cytochrome
oxydase de cette région corticale.
Par
conséquent, il a été suggéré que la ségrégation
entre les
signaux magnocellulaire et parvocellulaire persiste jusqu’aux niveaux
les plus élevés de l’analyse visuelle. La différence
fonctionnelle marquée entre la voie ventrale et la voie dorsale pourrait
d’ailleurs être attribuée à une contribution préférentielle
du canal P-IB pour la première et du canal M pour la seconde.
Pour les cellules de l’aire
MT, le déplacement d’un objet semble être tellement plus important
que la nature de cet objet que cette région du cortex est organisée
en colonnes codant pour l’orientation du mouvement, exactement comme les
colonnes d’orientation des lignes de V1.
Certaines cellules
de l’aire MT paraissent même répondre non pas à la direction
réelle, mais bien à la direction perçue du mouvement. Par
exemple deux groupes de lignes lumineuses se déplaçant chacun à
45 degrés de part et d’autre de la verticale donnent l'impression,
en se croisant, d'un mouvement vertical. Si ce sont les cellules ayant une préférence
pour un angle de 45 degrés qui vont répondre le mieux à ce
type de stimulus dans l’aire V1, plusieurs cellules de l’aire MT qui
présentent normalement une sélectivité de direction verticale
vont répondre de manière convaincante aux deux stimuli se déplaçant
à 45 degrés, donc à la direction apparente du mouvement.
Au-delà de l’aire MT se trouvent
d’autres régions impliquées dans l’analyse du mouvement
comme l’aire MST par exemple. Les cellules qui s’y
trouvent sont non seulement sensibles au déplacement linéaire comme
dans l’aire MT, mais également au déplacement radiaire (vers
un point ou à partir d’un point) ou encore au mouvement circulaire
(dans le sens horaire ou anti-horaire). Ils sont aussi sélectivement activés
par des configurations complexes de mouvements, correspondant au déplacement
des éléments de notre environnement quand on avance dans celui-ci.
Certains neurones de l'aire supérieure temporale polysensorielle (STP)
répondent même sélectivement à des mouvements biologiques
qui
ont pu s’avérer essentiels pour la survie de l’individu,
comme la reconnaissance de la démarche d'un congénère par
exemple.
Ces nombreuses données recueillies sur le mouvement
par la voie dorsale pourraient avoir plusieurs rôles : extraire de l’information
pertinente du flux d’objets qui traverse notre champ visuel quand nous nous
déplaçons pour guider nos déplacements; aider à l’orientation
du mouvement des yeux; ou encore identifier les objets qui se déplacent
autour de nous et qui ont une valeur de survie.
En plus du trajet séquentiel du signal
visuel à travers chaque niveau hiérarchique d’organisation
du système visuel, de nombreuses voies de traitement de l’information
fonctionnent aussi en parallèle, formant un réseau
de circuits d’une grande complexité. Cette complexité
n’est pas étrangère aux nombreuses boucles de rétroaction
que chacune de ces aires forme en retournant des connexions vers les régions
qui lui en ont envoyées. S’ajoutent également des projections
vers des structures sous-corticales comme le corps
genouillé latéral ou le colliculus
supérieur.