La lumière qui entre dans
l’oeil doit traverser toutes
les couches de la rétine avant d’atteindre
le segment externe des photorécepteurs. Le segment
interne des cônes, situé juste avant sur le
trajet du rayon lumineux menant aux pigments photosensibles,
jouerait un peu le rôle de fibres optiques pour guider
les rayons vers le segment externe. Ceci expliquerait pourquoi
les cônes sont beaucoup plus sensibles aux rayons lumineux
en provenance du centre de l’œil. Ces rayons sont
mieux alignés avec le segment interne et le traversent
plus efficacement que ceux qui l'atteignent obliquement.
Les cônes tirent ainsi partie de la meilleure optique
du centre de l’œil.
À l’opposé les bâtonnets, pour qui
chaque photon est important étant donné leur
rôle essentiel dans les conditions de lumière
réduite, ne peuvent se permettre de perdre des photons
et n’ont donc pas ce système de direction sélective
du segment interne des cônes.
De nouveaux disques sont constamment
produits à la base du segment externe des cônes
et des bâtonnets, de sorte que ceux-ci s’allongent
continuellement. C’est pourquoi l’épithélium
pigmenté qui est en contact avec eux phagocyte et détruit
les vieux disques qui se trouvent à l’extrémité
du segment externe.
LES PHOTORÉCEPTEURS
La transduction du signal
lumineux en influx nerveux est assumée par environ 125
millions de photorécepteurs qui sont situés dans
la partie la plus profonde de la
rétine. Deux
types de photorécepteurs bien différents se partagent
cette tâche. D'un côté les 120 millions
de bâtonnets qui
permettent de voir en nuances de gris dans des conditions de
faible luminosité
("la nuit, tous les chats sont gris"). Et de l'autre,
les 5 millions de cônes, plus petits
et plus larges, qui sont sensibles à la couleur dans
des conditions de forte luminosité.
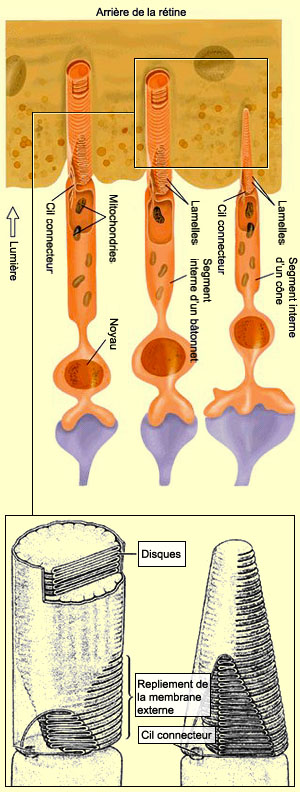
La forme du
segment externe, cylindrique pour les bâtonnets et cônique
pour les cônes, n’est pas la seule caractéristique
qui distingue les deux types de photorécepteurs. L’arrangement
et le nombre de disques à l’intérieur de
ce segment externe en est une autre. Celui des bâtonnets
est rempli d’environ 900 de ces disques qui sont complètement
indépendants du reste de la membrane et s’empilent
librement dans le segment externe. Les disques des cônes
sont beaucoup moins nombreux. Ils sont également formés
par le repliement de la membrane du segment externe, mais restent
cependant reliés à celle-ci contrairement à
ceux des bâtonnets.
Les photorécepteurs sont ni plus ni moins que des cellules
ciliées très spécialisées où les
segments externe et interne sont reliés par un cil connecteur.
Le segment interne des photorécepteurs contient le noyau
et les organites (les mitochondries, l'appareil de Golgi, etc.)
indispensables au fonctionnement de toute cellule.
Encore ici, des différences anatomiques notables sont
observées entre les segments internes des cônes
et des bâtonnets (voir encadré).
La
répartition des cônes et des bâtonnets varie
sur la surface rétinienne. Très peu
de cônes se retrouvent en périphérie
où les bâtonnets prédominent. À
l’opposé, il n’y a pas du tout de bâtonnets
dans la région centrale de la rétine appelée fovéa.
C’est pour cette raison que nos yeux bougent afin de
faire tomber l’objet d’intérêt du
champ visuel sur cette zone de grande acuité
visuelle.
Finalement, la distinction fonctionnelle
la plus importante entre cônes et bâtonnets, celle qui
rend les premiers sensibles à la couleur et pas les seconds,
vient de leurs photopigments différents. En effet, contrairement
aux bâtonnets qui ont toujours le même type de photopigment
appelé rhodopsine, le segment externe des cônes contient
l'une des trois différentes opsines absorbant d’une
manière maximales les ondes lumineuses à courte, moyenne
ou longue longueur d’onde. Ces trois pigments à la
sensibilité spectrale différente sont à la
base de notre
vision des couleurs.
La sensibilité de nos yeux à la
lumière n’est pas fixe mais s’adapte au
niveau de luminosité
de différentes façons. L’adaptation
à l’obscurité se produit par
exemple lorsque l’on entre dans une salle de cinéma alors
que la représentation est déjà
commencée. Nous avons d’abord l’impression
de ne voir absolument rien. Mais très rapidement nos iris
s’ouvrent pour laisser passer plus
de lumière vers la rétine. Un phénomène
plus lent se produit aussi au niveau du pigment photosensible
des bâtonnets, la
rhodopsine. Au cours des 20 ou 25 premières
minutes passées dans un environnement obscur, les stocks
de rhodopsine, épargnés par la lumière,
se régénèrent plus facilement de telle
sorte que la sensibilité des bâtonnets à
la lumière s’accroît d’environ un
million de fois !
Quand cet œil adapté à l’obscurité
retrouve la lumière, le phénomène inverse
se produit et l’on se retrouve, en sortant du cinéma,
temporairement aveuglé par le soleil. C’est l’adaptation
à la lumière dont la première
étape est la fermeture rapide de l’iris pour réduire
l’entrée de lumière dans l’œil.
Les autres modifications biochimiques s’inversent
également de sorte qu’après quelques minutes
seulement, notre vision s’est adaptée aux fortes
lumières d’une journée ensoleillée.
Les cônes s’adaptent donc plus rapidement à
la lumière que les bâtonnets à l’obcurité.
La fonction des photorécepteurs est
de transformer l’énergie lumineuse en potentiel
de membrane. La mécanique de ce processus est
comparable en plusieurs points à
celle que l’on retrouve au niveau des synapses
utilisant des récepteurs
métabotropes pour assurer la transduction
chimique. En effet, la fixation d'un neurotransmetteur
à un récepteur métabotrope active
des protéines G qui à leur tour stimulent
diverses enzymes. Ces enzymes vont modifier la concentration
intracellulaire de second messager, ce qui va amener
un changement dans la conductance de certains canaux
ioniques et par conséquent une variation du potentiel
membranaire.
La transduction de la lumière par les photorécepteurs
de la rétine suit les mêmes grandes étapes.
Mais avant de les mentionner, il faut d’abord souligner
le fait que, contrairement au potentiel de repos habituel
des neurones qui se situe autour de –
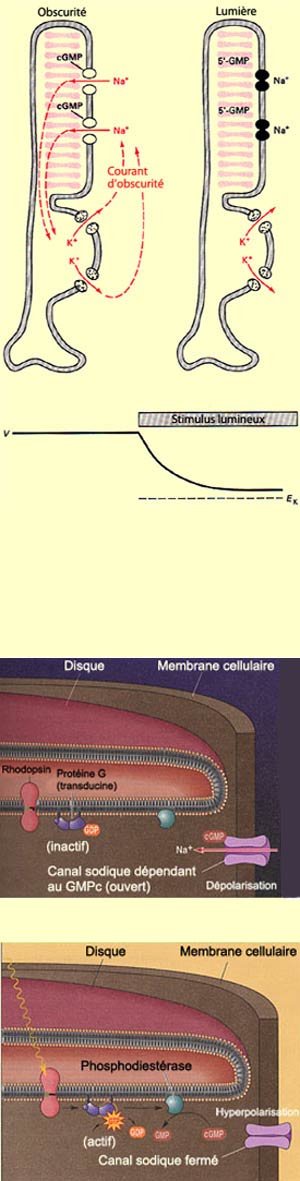
65 mV, le potentiel de membrane du segment externe des
bâtonnets est d’environ – 30 mV dans
l’obscurité. Cette dépolarisation
découle de l’entrée constante de
sodium par des canaux sodiques spécifiques du segment
externe en l’absence de lumière. Or
ces canaux sont maintenus ouverts par la présence
du second messager GMPc (guanosine monophosphate cyclique)
produit en continu par l’enzyme guanylate cyclase
en condition d’obscurité. Ce phénomène
est appelé le courant d’obscurité.
Quand des photons lumineux frappent le pigment photosensible
des cônes ou des bâtonnets, celui-ci
change de conformation et active ainsi une protéine
G. Appelée transducine, cette protéine
G va faire diminuer le taux de GMPc dans le photorécepteur
en activant l'enzyme phosphodiestérase qui détruit
le GMPc. Cette baisse de GMPc réduit à
son tour la conductance au sodium et, par conséquent,
le courant d’obscurité responsable du potentiel
de membrane élevé du segment externe.
Le résultat est donc le contraire de ce que l’on
pourrait s’attendre, à savoir que la présence
de lumière va hyperpolariser le photorécepteur.
Et par conséquent faire en sorte que moins de
neurotransmetteurs vont être relâchés
à la synapse avec les
cellules bipolaires.