Les photorécepteurs de la rétine des vertébrés,
les bâtonnets et les cônes, se
distinguent par de nombreuses caractéristiques,
tant anatomiques que fonctionnelles. La principale étant le rôle opposé que
jouent les deux types de photorécepteurs : bâtonnets
très sensibles dans les basses intensités lumineuses
mais ne distinguant pas les couleurs (vision scotopique); et cônes
nécessitant de fortes lumières mais permettant une
vision précise et en couleur du monde qui nous entoure
(vision photopique).
Mais dans les deux cas, c’est l’absorption de l’énergie
lumineuse par les photopigments fichés dans les disques du segment externe
qui initie la réponse nerveuse, en l’occurrence
une hyperpolarisation
de la cellule photoréceptrice. Dans le cas des
bâtonnets, ce
pigment photosensible s’appelle la rhodopsine et a son pic de sensibilité autour
de 500 nanomètres (nm) dans le spectre électromagnétique
de la lumière visible.
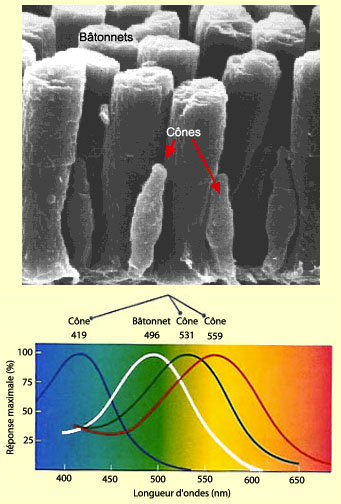
Les cônes
contiennent pour leur part trois variétés d’opsine, une
protéine transmembranaire très proche de la rhodopsine.
Des différences dans la séquence des acides aminés
de cette protéine rendent compte de leur courbe d’absorption
différente de la lumière (voir le graphique).
Par conséquent, ces trois variétés d’opsine
déterminent trois types de cônes à la sensibilité
spectrale différente.
Les cônes « bleus » sont principalement activés
par une longueur d’onde d’environ 420 nm, les cônes « verts » autour
de 530 nm, et les cônes « rouges » près
de 560 nm. Alternativement, on parle aussi de cônes-S
pour décrire les cônes qui contiennent en majorité le
pigment sensible au bleu; de cônes-M pour ceux qui présentent
une concentration plus importante en pigments sensibles au
vert; et de cônes-L pour les cônes porteurs du
pigment sensible au rouge.
Les trois pigments sont donc présents dans chaque type
de cône, mais en proportion très majoritaire pour
la couleur dominante. Les lettres S, M ou L viennent de l’anglais « Short,
Medium and Long wavelenght
» qui désigne les longueurs d’onde courtes,
moyennes et longues d’absorption maximale des différentes
opsines.
Un objet dont la couleur se situe quelque part dans le spectre visible va donc
exciter à divers degrés les 3 types de cône. Un objet vert
par exemple va surtout stimuler les cônes verts, mais aussi les rouges à
un moindre degré et très légèrement
les bleus. Notre perception des couleurs dépend donc
de cette superposition des différents spectres d’absorption
des trois types de cônes. Et bien sûr, par la
suite, de la
complexité des interactions neuronales dans la rétine le
reste du cerveau.
Le daltonisme est
une déficience de la vision qui se caractérise
par une incapacité à différencier certaines
teintes ou couleurs. Le nom de cette déficience vient
de celui du physicien John Dalton (1766-1844) qui en était
lui-même atteint.
Environ 8 % des hommes en sont atteints à divers degré
et un peu moins de 1 % des femmes. Cette différence
s’explique par le fait que la principale forme de daltonisme
est héréditaire et que les mutations génétiques
qui en sont la cause surviennent sur le chromosome X. Comme
le gène muté est récessif, les femmes
qui possèdent deux exemplaires du chromosome X peuvent
être porteuses sans être atteintes si l’autre
chromosome X est intact. Mais pas les hommes, qui n’en
possèdent qu’un, et qui sont donc automatiquement
atteint s’ils ont la mutation.
Les cas de daltonisme total, l'achromatopsie,
où le monde n’apparaît qu’en teinte
de gris sont très rares. Plus souvent, les daltoniens
ont de la difficulté à distinguer entre le rouge
et le vert et, beaucoup plus rarement, entre le bleu et le
jaune.
Le daltonisme classique au rouge et au vert est le résultat,
au niveau de la rétine, d’un manque de cônes
« rouges ». On classe en général
les formes de daltonisme selon le type du cône atteint.
Aux trois types de cônes correspondent donc trois types
de daltonismes, où la deutéranopie, la déficience
du pigment vert, est la plus fréquente.
L’adaptation à l’obscurité
se fait en deux temps qui reflètent la transition d’une
vision issue des cônes (vision photopique) à
une vision issue des bâtonnets (vision scotopique). Durant
les 5 ou 6 premières minutes après le passage
d’une pièce éclairée à
une pièce sombre, le seuil de sensibilité à
la lumière, à l’origine très élevé
à cause du séjour dans la pièce éclairée,
décroît d’abord rapidement pour ensuite
sembler vouloir se stabiliser de manière asymptotique.
Mais autour de la septième minute, le seuil commence
à décliner de plus belle et atteint un second
plancher asymptotique, beaucoup plus bas celui-là, environ
une demi-heure plus tard. Ce seuil minimal est celui de la
vision scotopique alors que le seuil initial représente
celui de la vision photopique.
LA TRANSDUCTION
DU SIGNAL LUMINEUX
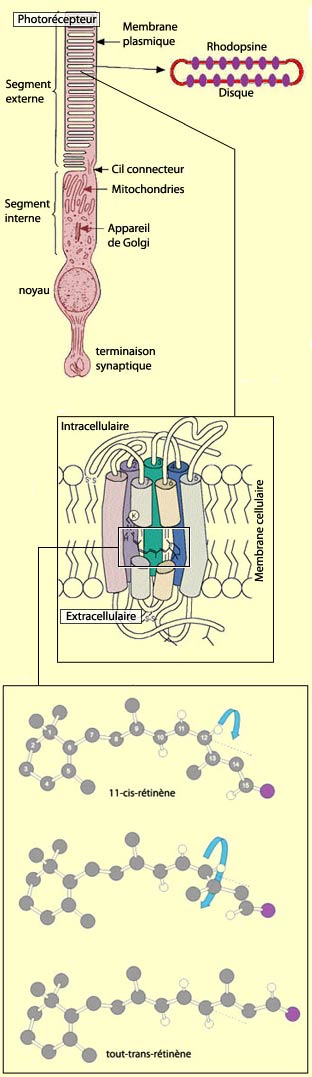
La transduction
de l'énergie lumineuse en variation de potentiel
de membrane des photorécepteurs commence avec l’absorption
de photons par un pigment photosensible appelé opsine.
Dans le cas des bâtonnets, il s’agit d’une
protéine appelée rhodopsine,
et dans le cas des cônes, des iodopsines.
Un seul bâtonnet peut compter jusqu’à
100 millions de molécule de rhodopsine sur les disques
de son segment externe. Ces molécules de la famille
des opsines comprennent sept domaines transmembranaires.
La structure de cette protéine n’est pas sans
rappeler celle
des récepteurs synaptiques métabotropes.
On peut d’ailleurs considérer la rhodopsine
comme une protéine-récepteur qui serait déjà associée à son
agoniste. En effet, une molécule dérivée
de la vitamine A appelée rétinène (ou
rétinal) est attaché au milieu du septième
domaine transmembranaire de la rhodopsine. Plus précisément,
il s’agit de la forme « 11-cis » du rétinène
qui, lorsqu’elle absorbe la lumière, subit un
changement de conformation vers la forme «
tout-trans ». C’est ce changement de forme, appelé aussi
isomérisation, qui fait passer la rhodopsine à sa
forme active, la metarhodopsine II. Le dépliement
du rétinène produit donc le même effet
que si un neurotransmetteur venait soudainement de se fixer à son
récepteur.
La metarhodopsine II va par la suite stimuler une protéine
G particulière appelé transducine. Comme
toutes les protéines G, celle-ci va rendre active
une autre enzyme qui est dans ce cas-ci la phosphodiestérase
(PDE). Quand la phosphodiestérase est activée,
elle transforme le GMPc présent en l’absence
de lumière dans le cytoplasme des bâtonnets
en simple GMP. Et c’est cette chute du taux de GMPc
qui entraîne la fermeture des canaux sodiques et
l’hyperpolarisation de la membrane. Résultat
: moins de neurotransmetteurs (probablement du glutamate)
relâchés aux terminaisons synaptiques des
photorécepteurs.
À noter l’amplification du signal qui survient
à deux endroits dans la cascade biochimique : chaque
metarhodopsine II active autour d’une centaine de
protéines G, et chaque molécule de phosphodiestérase
va permettre l’hydrolyse d’environ un millier
de molécules de GMPc en GMP. C’est ce phénomène
d’amplification qui permet aux bâtonnets de
détecter la présence d’un seul photon
lumineux.
La vitamine A produite à partir
de la bêta-carotène de plusieurs de nos aliments
(dont évidemment la carotte) est nécessaire
pour la synthèse du rétinène situé
au centre de la molécule de rhodopsine. En effet,
une déficience sévère en vitamine A
amène une altération de la vision en basse
luminosité
due à la faible quantité de rétinène
produit. Durant la journée cependant, la quantité
de lumière est généralement suffisante
pour permettre une vision relativement normale malgré
le bas niveau de pigments visuels.