Intuitivement, on pourrait s’attendre à
ce que la couche de photorécepteurs se retrouve la première à
partir de l’intérieur du globe oculaire pour recevoir la lumière
en premier. Mais ce n’est pas ce que l’on observe: avant d’atteindre
les photorécepteurs, la lumière doit plutôt traverser l’ensemble
des différentes couches formant la rétine.
Cet arrangement
de prime abord un peu paradoxal s’explique par le fait que le pigment photosensible
contenu dans les photorécepteurs doit être en contact avec la couche
de cellules épithéliales du fond de l’œil qui lui fourni
un apport continuel en rétinène,
le dérivé de la vitamine A sensible à la lumière.
Une fois leur conformation changée par l’énergie lumineuse,
les molécules de rétinène sont également recyclées
dans cet épithélium. La pigmentation foncée de celui-ci empêche
aussi les photons non absorbés d’être réfléchis
à nouveau vers les photorécepteurs et ainsi de créer une
lumière parasite qui brouillerait l’image.
Les cellules horizontales partagent avec les
cellules amacrines une caractérisque particulière : l’absence
d’un quelconque prolongement qui pourrait ressembler à un axone.
Ces cellules ne possèdent en fait que des dendrites dont certaines peuvent
être présynaptiques, autrement dit jouer le rôle d’un
axone. Les prolongements de ces cellules jouent donc apparemment les deux rôles.
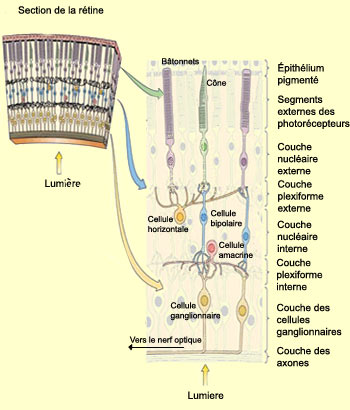
LA RÉTINE
Dans la rétine,
l’information voyage des photorécepteurs aux cellules bipolaires
puis aux cellules ganglionnaires. À chaque relais de cette
voie la plus directe, les réponses sont modifiées par l’activation
de connexions latérales grâce aux cellules horizontales et amacrines.
Le début de l’analyse des stimuli visuels commence donc à
même la rétine.
Curieusement, la
lumière doit traverser successivement les différentes couches cellulaires
de la rétine avant d’atteindre les photorécepteurs (voir encadré).
La
première est la couche des cellules ganglionnaires, formée
des corps cellulaires des cellules du même nom. Elle est suivie par la couche
plexiforme interne, un enchevêtrement d’axones et de dendrites
provenant des cellules ganglionnaires, des neurones bipolaires et des cellules
amacrines. Puis vient la couche nucléaire interne, formée
des corps des cellules bipolaires, horizontales et amacrines. On retrouve ensuite
la couche plexiforme externe composée des terminaisons
nerveuses des cellules bipolaires, horizontales et des photorécepteurs.
Vient ensuite la couche nucléaire externe, celle des corps
cellulaires des photorécepteurs. Finalement la couche des segments
externes des photorécepteurs qui contient les pigments
sensibles à la lumière. Et ce sont les extrémités
de ces segments externes qui sont enchâssés dans l’épithélium
pigmenté.
Chacun des types cellulaires
de ces couches possède une répartition et des caractéristiques
physiologiques qui leur sont propres.
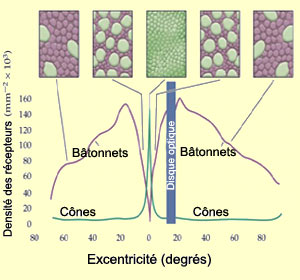
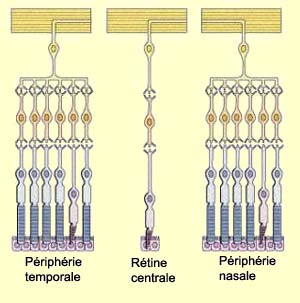
La répartition des cônes
et des bâtonnets n’est pas uniforme sur la rétine,
les bâtonnets étant beaucoup plus nombreux dans la rétine
périphérique et les cônes dans la
fovéa. Le nombre de photorécepteurs connectés à
une même cellule ganglionnaire est aussi beaucoup plus grand en périphérie.
L’effet combiné de cette organisation est d’accroître
la sensibilité à la lumière en périphérie de
la rétine. La contrepartie est que la précision de l’image
souffre de la convergence de nombreux photorécepteurs sur une même
cellule ganglionnaire.
Une
bonne acuité visuelle comme celle de la rétine centrale demande
en effet un faible rapport photorécepteurs/cellules ganglionnaires. Elle
est aussi favorisée par les cônes de la fovéa qui sont très
petits et tassés les uns contre les autres. Plus on s’éloigne
de la fovéa, plus la taille des cônes augmente ainsi que l’espace
entre eux, les bâtonnets remplissant l’espace restant. Malgré
la grande densité des cônes à la fovéa, la petitesse
de cette région fait en sorte que seulement 1% des cônes de la rétine
s’y trouvent.
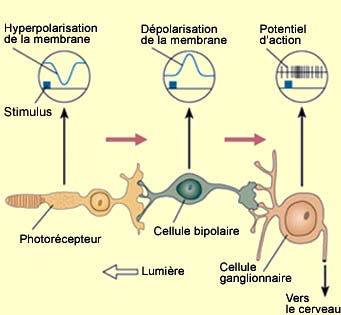
Les neurones
bipolaires sont un passage obligé entre les photorécepteurs
et les cellules ganglionnaires, tant pour la voie directe que pour la voie indirecte
faisant intervenir les cellules horizontales. Le signal s’y transmet sous
forme de potentiel gradué. Celui-ci peut être autant une dépolarisation
qu’une hyperpolarisation, selon
le type ON ou OFF de la cellule bipolaire.
Une cellule bipolaire reçoit
des connexions synaptiques directes d’un certain nombre de photorécepteurs
situés plus ou moins vis-à-vis elle: de un au centre de la fovéa,
jusqu’à des milliers dans la rétine périphérique.
En plus de ces connexions directes avec les photorécepteurs,
les cellules bipolaires reçoivent des afférences de cellules horizontales.
Celles-ci sont reliées à un ensemble de photorécepteurs qui
entourent le groupe central faisant les connexions directes aux cellules bipolaires.
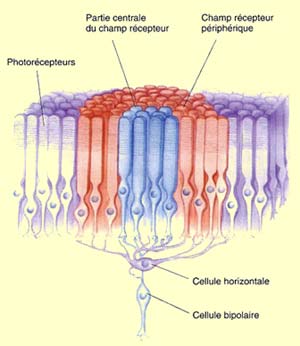
Par conséquent, le
champ récepteur des cellules bipolaires comprend deux parties : un
champ récepteur central constitué de l’information qui transige
directement des photorécepteurs aux cellules bipolaires; et un champ récepteur
périphérique qui reçoit l’information passant par les
cellules horizontales.
Comme les cellules horizontales
sont connectées latéralement à plusieurs cônes, bâtonnets
et neurones bipolaires, leur rôle est d’inhiber l’activité
des cellules avoisinantes. Cette suppression sélective de certains signaux
nerveux s’appelle l’inhibition latérale et son rôle général
est d’augmenter l’acuité d’un signal sensoriel. Dans
le cas de la vision, quand une source lumineuse atteint la rétine, elle
peut illuminer fortement certains photorécepteurs et d’autres beaucoup
moins. En supprimant le signal de ces photorécepteurs moins illuminés,
les cellules horizontales assurent que seul le signal des photorécepteurs
bien illuminés est transmis aux cellules ganglionnaires, améliorant
ainsi le contraste et la définition du stimulus visuel.
La morphologie
des cellules amacrines est très diversifiée et
elles utilisent un nombre impressionnant de neurotransmetteurs. Leurs corps cellulaires
sont tous situés dans la
couche nucléaire interne et leurs terminaisons synaptiques dans la
couche plexiforme interne. En reliant les neurones bipolaires et ganglionnaires,
elles forment une route alternative indirecte entre ceux-ci. Les cellules amacrines
semblent avoir plusieurs fonctions, la plupart encore inconnues.
Les circuits formés par les cellules amacrines
dans la couche plexiforme interne apportent de l’information supplémentaire
aux cellules ganglionnaires, possiblement en augmentant encore davantage le contraste
entre le centre et la périphérie généré grâce
aux cellules horizontales.
La nature excitatrice ou inhibitrice de la synapse
photorécepteur-cellule bipolaire peut dépendre soit du type de neurotransmetteur
relâché par les photorécepteurs, soit du type de récepteurs
sur la membrane post-synaptique de la cellule bipolaire. On croit de moins en
moins à la possibilité qu’un photorécepteur puisse
relâcher deux neurotransmetteurs différents et tout porte à
croire que les cellules bipolaires ON et OFF ont différents récepteurs
moléculaires.
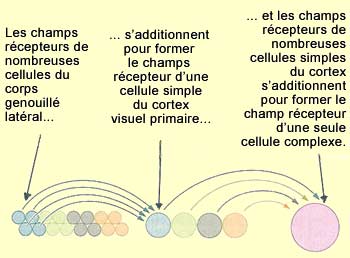
LES CHAMPS RÉCEPTEURS, DE LA RÉTINE AU CORTEX
Les neurones bipolaires ont
un champ
récepteur de type centre-périphérie. Le centre du champ
est alimenté par les connexions directes d’un petit nombre de photorécepteurs
tandis qu’à la périphérie les inputs proviennent d’un
ensemble plus large de photorécepteurs dont l’activité est
recueillie par les
cellules horizontales.
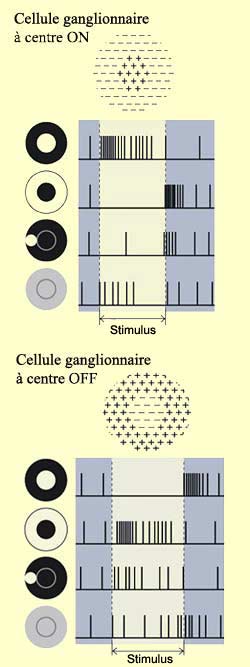
L’éclairement
du centre d’un champ récepteur d’une cellule bipolaire produit
un changement opposé à l’éclairement de sa périphérie
sur le
potentiel de membrane de la cellule bipolaire. Dans le cas du schéma
ci-contre par exemple, pour certaines cellules bipolaires dites à centre
ON, l’éclairement du centre entraîne d’abord une hyperpolarisation
du photorécepteur, hyperpolarisation qui se transforme en dépolarisation
dans le neurone bipolaire à
cause du caractère inhibiteur de la synapse qui les relie. Le résultat
sur le neurone ganglionnaire qui suit est une excitation qui se traduit par une
augmentation de la fréquence des potentiels d’action qu’il
émet.
Source
: Adapté de J.E. Dowling
Par
ailleurs, la même cellule bipolaire à centre ON subirait une hyperpolarisation
si la lumière frappait plutôt la périphérie de son
champ récepteur. Et à l’inverse, d’autres neurones bipolaires
subissent une dépolarisation quand c’est une zone d’obscurité
qui frappe le centre de leur champ récepteur et une hyperpolarisation quand
cette zone atteint la périphérie. On parle alors de neurones bipolaires
à centre OFF.
Cette
organisation centre-périphérie des champs récepteurs des
cellules bipolaires va se transmettre aux cellules ganglionnaires par des synapses
situées dans la couche plexiforme
interne.
Ainsi, des synapses relient les cellules bipolaires à
centre ON aux cellules ganglionnaires à centre ON, et les cellules bipolaires
à centre OFF aux cellules ganglionnaires à centre OFF. L’accentuation
des contrastes générée par les champs récepteurs centre-périphérie
des cellules bipolaires est donc préservée et transmise aux cellules
ganglionnaires, puis jusqu’au cortex.
Car notre vision dépend
en grande partie de notre capacité à discerner les contrastes entre
un objet et le fond devant lequel il se trouve. L’établissement de
voies
de traitement de l’information visuelle parallèles dès
la rétine est un des moyens qui permet cette discrimination.
En plus des cellules
simples qui se rencontrent principalement dans le couche IV du cortex visuel,
on trouve en dehors de cette couche IV des cellules qui ne répondent que
si un stimulus lumineux présente une orientation donnée et est en
mouvement.
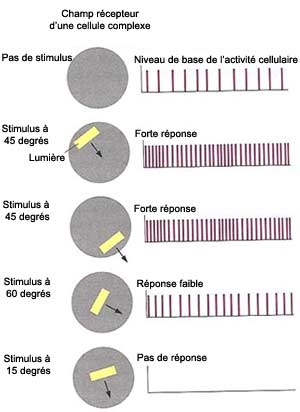
On appelle ces cellules les cellules
complexes. Elles peuvent détecter le mouvement d’abord à
cause de la convergence qui s’exerce sur elles par les axones de plusieurs
cellules simples de même orientation et de champs visuels voisins; et d’autre
part, grâce au phénomène de sommation temporelle : si une
cellule est excitée à nouveau peu de temps après une excitation
précédente, la membrane est encore assez dépolarisée
pour pouvoir à nouveau atteindre le seuil de déclenchement des potentiels
d’action avec un stimulus normalement insuffisant. Le déplacement
d’un trait de lumière active donc successivement plusieurs cellules
simples dont la sommation temporelle des stimulations fait répondre la
cellule complexe au mouvement.
Les cellules complexes montrent aussi
fréquemment une sélectivité à la direction, répondant
seulement quand le stimulus bouge dans une direction et pas dans l’autre.
Et contrairement aux cellules simples, elles ne sont pas tatillonnes sur l’endroit
où se trouve la bande de lumière dans leur champ. Les cellules complexes
constituent un nouveau niveau de traitement de l'information, mais certainement
pas le dernier puisqu’on a aussi découvert des cellules
hypercomplexes.
Comme dans les autres relais des voies visuelles,
il y a beaucoup plus de neurones corticaux recevant de l’information de
la partie centrale de la rétine que de ses régions périphériques.
Cette « rétinotopie » reflète un principe que
l’on retrouve dans d’autres régions du cortex et qui veut qu’une
plus grande précision sensorielle ou motrice
nécessite l’implication d’une plus grande surface corticale.
L'ORGANISATION
CELLULAIRE DU CORTEX VISUEL
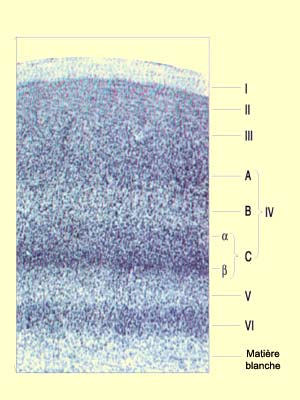
Le
cortex visuel primaire, comme toutes les aires du néocortex,
possède une organisation cellulaire stratifiée. Les couches I à
VI décrites originellement par Brodmann ont dues être subdivisées
à leur tour à mesure que se précisaient les voies d’entrée
et de sortie du cortex visuel.
C’est ainsi que la couche IV a été
subdivisée en trois sous-couches nommées IV A, IV B et IV C. La
couche IV C a elle-même été subdivisée à son
tour en IV C alpha et IV C bêta quand on a pu mettre en évidence
une différence dans la connectivité des cellules de la partie supérieure
et de celles de la partie inférieure de cette couche.
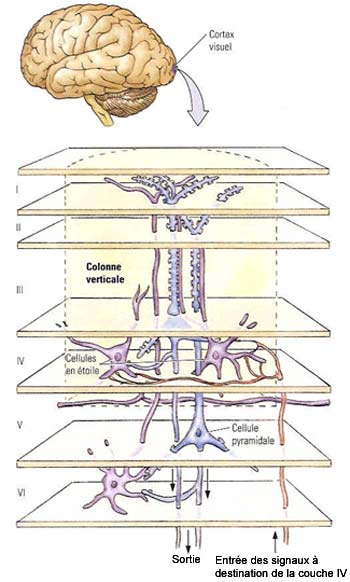
Les
axones de cellules du corps genouillé latéral transmettent l’information
en provenance de l’œil selon différentes voies qui se projettent
principalement dans la couche IV C. De plus, les cellules voisines de cette couche
IV C reçoivent des informations issues de régions voisines de la
rétine, préservant ainsi une organisation rétinotopique.
On sait par ailleurs que les informations issues du corps genouillé latéral
utilisent
des canaux séparés découlant de l’organisation
interne de cette structure.
Les axones des cellules
étoilées de la couche IV C qui reçoivent ces afférences
vont rejoindre à leur tour les dendrites des cellules pyramidales des couches
IV B et III. Ces cellules pyramidales projettent ensuite leur axone vers d’autres
aires corticales. Quant aux autres voies de sortie du cortex visuel primaire,
on sait que les cellules pyramidales de la couche V projettent au niveau sous-cortical
vers le collicule supérieur et la protubérance et que les axones
de la couche VI retournent massivement vers le corps genouillé latéral,
exerçant ainsi une rétroaction sur cette structure.
Si cette
stratification horizontale en couches est facilement visualisable avec des colorations
simples des neurones corticaux, une autre organisation, verticale celle-là,
n’a pu être mise en évidence que par des enregistrements électrophysiologiques
dans ces neurones.
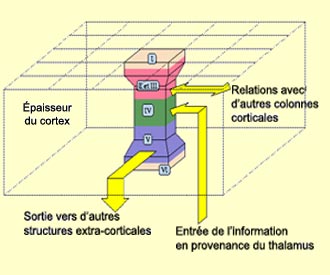
Les
premiers à avoir proposé cette organisation « en colonne »
qui se superpose à l’organisation en couche sont David Hubel et Torsten
Wiesel. Explorant les champs récepteurs des neurones du cortex visuel à
l’aide de microélectrodes, ils montrèrent que le cortex visuel
peut être considéré comme la juxtaposition de colonnes sensiblement
identiques. D'une colonne à l'autre, la différence vient simplement
de la portion du champ visuel qui leur est affectée. Les fonctions étagées
des différentes couches dans l'épaisseur de la colonne restent les
mêmes, mais chaque
colonne s’occupe de traiter une caractéristique d’une région
donnée du champ visuel (le contraste, la couleur, l’orientation,
le mouvements, etc.).