Six muscles oculomoteurs sont
fixés sur le globe oculaire et nous permettent de faire différents
types de mouvement avec nos yeux. Certains sont lents comme lorsque que l’on
suit du regard un objet qui se déplace devant nous. D’autres, appelés
saccades, sont très rapides. Ce sont eux qui font que vos yeux, qui doivent
demeurer fixes un instant pour analyser l’image, sautent très rapidement
d’un mot à l’autre pendant que vous êtes en train de
lire ces lignes par exemple.
L'OEIL
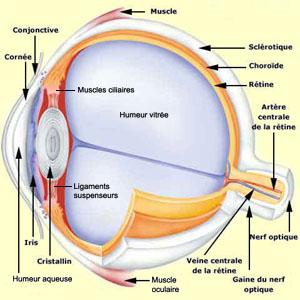
Une coupe transversale de l’œil
révèle que la première membrane traversée par la lumière
est la conjonctive. Il s’agit d’une fine membrane
transparente qui couvre le devant de l’œil et se replie pour tapisser
l’intérieur des paupières.
La cornée est en continuité avec le blanc de l’œil,
ou sclérotique, qui forme la paroi dure du globe oculaire
et dans laquelle sont insérées trois paires de muscles. Ce sont
ces muscles oculaires qui permettent les mouvements du globe
oculaire dans les orbites du crâne.
Située entre la sclérotique
et la rétine, la choroïde est une couche richement
vascularisée qui assure la nutrition de l'iris et de la rétine.
Elle contient de nombreux pigments foncés qui font que l'intérieur
de notre œil, visible à travers la pupille, est noir
L’humeur
aqueuse et l’humeur vitrée jouent un rôle fondamental dans
la focalisation de l’image sur la rétine grâce au phénomène
de réfraction. Ce phénomène se produit quand la lumière
passe dans un milieu où sa vitesse se trouve réduite, comme c’est
le cas dans les deux humeurs de l’œil par rapport à l’air.
Le rayon lumineux est alors dévié selon une ligne perpendiculaire
à la surface de partage des milieux.
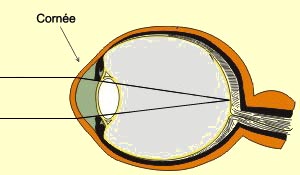
La courbure de la cornée
accentue aussi la réfraction des rayons lumineux virtuellement parallèles
provenant d’objets très éloignés. Les rayons qui arrivent
au centre de la cornée la frappent perpendiculairement et, n’étant
pas déviés, continuent tout droit jusqu’au centre de la rétine.
Les autres rayons qui arrivent sur la courbure de la cornée seront déviés
vers l’intérieur et arriveront exactement sur le même point
central de la rétine pour former une image au foyer chez la personne qui
n’a pas de troubles
de la vision.
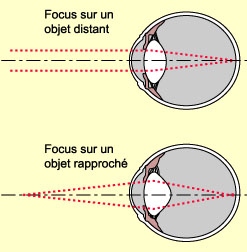
Si la cornée est responsable de la majeure partie
de la réfraction de la lumière dans l’œil, le cristallin
contribue également, mais dans une moindre mesure, à réfracter
les rayons lumineux venant de loin pour qu’ils convergent en un seul point
sur la rétine. Mais à plus courte distance, à partir de 9
mètres et moins environ, le cristallin joue un rôle beaucoup plus
actif pour nous aider à faire la mise au point. Car dans la formation de
l’image des objets plus rapprochés, les rayons qui parviennent à
l’œil sont plutôt divergents et la réfraction doit être
plus forte pour les faire converger sur la rétine. Cette mise au point
supplémentaire se fait par la modulation de la forme du cristallin.
En effet, au cristallin s’attachent
des ligaments suspenseurs qui le lient aux muscles ciliaires,
eux-mêmes attachés à la sclérotique. Comme ces muscles
forment un anneau à l’intérieur de l’œil, quand
ils sont relâchés, la tension sur les ligaments suspenseurs est forte
ce qui rend le cristallin plus plat et donc moins convergent. À l’opposé,
la contraction des muscles ciliaires diminue la tension qu’ils exercent
sur le cristallin. Ce dernier, tend alors à retrouver sa forme naturelle
et devient plus bombé, donc plus convergent pour les rayons lumineux. La
puissance de réfraction accrue qu’acquiert ainsi le cristallin permet
la formation d’une image nette des objets rapprochés sur la rétine,
un phénomène connu sous le nom d’accommodation.
Malgré la séparation des fibres
nerveuses issues de l’œil gauche et de l’œil droit dans
les différentes couches du CGL, chacune de celles-ci présente une
organisation rétinotopique
(présente aussi dans le colliculus
supérieur). De plus, les cartes des différentes couches
coïncident, de sorte qu’un axe perpendiculaire aux six couches du CGL
passe par des neurones sensibles aux mêmes points du champ visuel binoculaire,
donc perçus simultanément par l’œil droit et par l’œil
gauche.
LES CIBLES DU NERF OPTIQUE
Les axones des cellules
ganglionnaires parcourent le
nerf optique, le chiasma optique et le tractus optique. Ils contournent le
mésencéphale, cheminent sur la face médiale du lobe temporal
et se terminent, pour 80% d’entre eux, dans le corps genouillé
latéral (CGL). Le CGL, situé dans la partie dorsale du
thalamus, constitue donc la cible majeure de chaque tractus optique.
En coupe transversale, les CGL gauche
et droit montrent une organisation en six couches de cellules distinctes. Quand
on les considère dans les trois dimensions, les six couches du CGL ressemblent
à une pile de crêpes superposées qui se replient autour du
tractus optique comme l’articulation du genou, d’où leur nom.
La distribution des neurones du CGL en différentes couches indique
que des aspects distincts de l’information visuelle en provenance de la
rétine pourraient être traités séparément au
niveau de ce relais synaptique. Et c’est exactement ce que l’on a
pu démontrer.
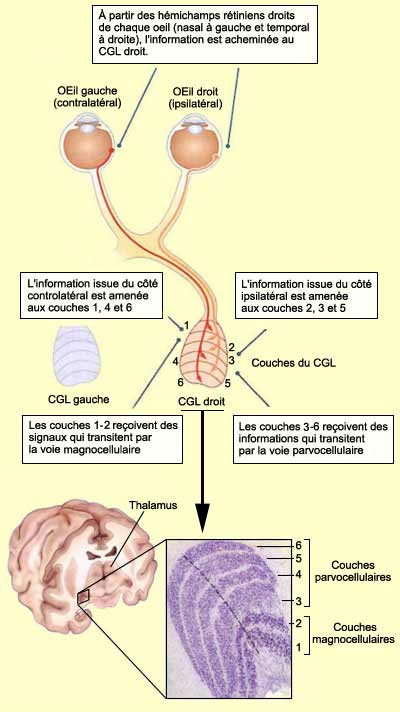
En effet, comme le CGL droit traite tous les signaux
visuels en provenance du champ visuel gauche (et l’inverse pour le CGL gauche),
il reçoit des axones de la rétine nasale gauche et de la rétine
temporale droite. Or on observe que les axones issus de l’œil ipsilatéral
(ici, le droit) forment des synapses sur les couches cellulaires 2, 3 et 5 du
CGL tandis que ceux issus de l’œil controlatéral (ici, le gauche)
se projettent au niveau des couches 1, 4 et 6.
On peut aussi observer que les couches ventrales
1 et 2 contiennent des plus gros neurones que les couches plus dorsales 3, 4,
5 et 6. On utilise donc le terme magnocellulaire (M) pour désigner
les deux couches ventrales et celui de parvocellulaire (P) pour
parler des quatre autres, exactement comme on le fait pour les
cellules ganglionnaires de type M et de type P. En fait, on a pu démontrer
que ce sont très exactement les cellules ganglionnaires de type M qui projettent
dans les couches magnocellulaires du CGL et les cellules ganglionnaires de type
P dans les couches parvocellulaires. Le traitement en parallèle de canaux
d’information distincts à partir de la rétine semble donc
être préservé à travers le CGL.
Même
les petits neurones situés sur la partie ventrale de chaque couche du CGL
qui forment les couches coniocellulaires reçoivent des
connexions des cellules ganglionnaires non M-non P de la rétine, ce qui
confirme la ségrégation de l’information en provenance des
différents types de cellules ganglionnaires.
Malgré la
grande influence qu’a l’innervation en provenance de la rétine
sur l’organisation du CGL, environ 80 % des connexions excitatrices qui
entrent dans le CGL ne proviennent pas de la rétine mais bien du cortex
visuel primaire et d'autres structures sous-corticales ! Il semble donc que le
cortex visuel primaire exerce une rétroaction importante sur le CGL.
En d’autres termes, la cible principale du CGL pourrait modifier en retour
les réponses visuelles de ce même CGL.
Une
autre observation renforce l’idée que le CGL, tout comme les
autres structures sous-corticales impliquées dans la vision, n’est
pas un simple relais passif sur la voie qui va de la rétine au cortex :
son activation possible par des neurones du tronc cérébral dont
l’activité est associée à la vigilance et aux processus
attentionnels. Ceux-ci agiraient comme modulateur de la réponse des neurones
du CGL, renforçant l’idée que le CGL est en réalité
le premier endroit de la voie visuelle où des états mentaux particuliers
influencent notre perception visuelle.
Contrairement aux daltoniens
qui confondent certaines couleurs, il existe de rares cas de personnes qui ne
voient aucune couleur. On utilise le terme d’achromatopsie
pour désigner cette incapacité.
L’achromatopsie
peut être congénitale, comme lors d’une déficience génétique
au niveau des cônes de la rétine, ou elle peut être acquise,
à la suite par exemple d’un accident cérébrovasculaire.
Les études d’imagerie cérébrale chez les personnes
souffrant d’achromatopsie acquise ont confirmé des lésions
importantes à l’aire V4 du cortex visuel, région reconnue
pour le traitement des couleurs. Ces personnes décrivent leur perception
visuelle du monde comme terne et grise, un peu comme sur une vieille télévision
noir et blanc.
L’achromatopsie est aussi généralement
associée à une certaine altération de la reconnaissance de
la forme des objets, ce qui est cohérent avec le rôle de reconnaissance
de la forme des objets qu’a également le système ventral.
L’organisation
hiérarchique entre un cortex primaire et des cortex secondaires se retrouve
à plusieurs endroits dans le cerveau, que ce soit dans le cortex sensoriel,
moteur
ou auditif. Les aires secondaires de ces différents cortex convergent ensuite
vers des zones dites «associatives». Celles-ci ont un niveau de traitement
plus global de l’information. Elles associent progressivement des signaux
en provenance d’autres modalités sensorielles pour faire émerger
une représentation intégrée et multisensorielle du monde.
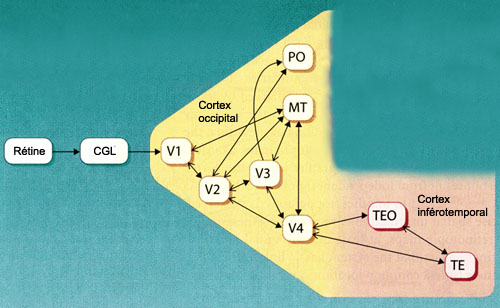
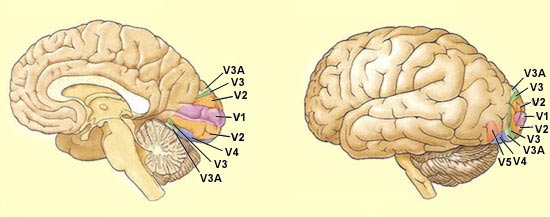
On
a découvert jusqu’à ce jour près d’une trentaine
d’aires corticales différentes qui contribuent à la perception
visuelle. Les aires
primaires (V1) et secondaires (V2) sont entourées de nombreuses autres
aires visuelles tertiaires ou associatives : V3, V4, V5 (ou MT), PO, etc.
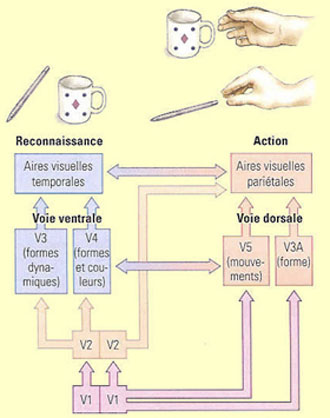
Un
schéma général émerge toutefois de cette complexité
selon lequel il existerait deux grands systèmes corticaux de traitement
de l’information visuelle : une voie ventrale qui s’étendrait
vers le lobe temporal, et une voie dorsale qui se projette vers le lobe pariétal.
La voie
ventrale aurait pour mission fondamentale de permettre la perception
consciente, la reconnaissance et l'identification des objets en traitant leurs
propriétés visuelles "intrinsèques" comme leur
forme, leur couleur, etc. La
voie dorsale, en revanche, aurait pour mission fondamentale d'assurer le contrôle
visuo-moteur sur les objets en traitant leurs propriétés "extrinsèques",
celles qui sont critiques pour leur saisie, comme leur position spatiale, leur
orientation ou leur taille.
Dans les aires corticales contribuant
au système ventral, des représentations de plus en plus
complexes et spécialisées du monde extérieur s’élaborent.
L’aire V3 reçoit
d’importantes connexions de l’aire V2 et envoie des projections vers
les aires MT et V4. Plusieurs des neurones de l’aire V3 ont des propriétés
semblables à l’aire V2, étant par exemple pour la plupart
sélectif à l’orientation. Mais l’aire V3, encore mal
connue, possède aussi des cellules qui ont des propriétés
plus complexes. Des cellules qui sont par exemple sensibles à la couleur
et au mouvement, caractéristiques que l’on retrouve davantage dans
les stades subséquents de l’analyse du signal visuel.
Après
être passée par les aires V1, V2 et V3, une partie de l’information
visuelle chemine ventralement vers le cortex temporal en passant tout d’abord
par l’aire V4. Celle-ci reçoit de l’information
de la
région des taches et des zones intertaches du cortex strié,
via un relais dans V2. Comme c’est le cas dans les aires visuelles autres
que V1 (on dit aussi « extrastriées »), les cellules de l’aire
V4 présentent des champs récepteurs plus larges que ceux du cortex
strié. De plus, les champs récepteurs de V4 sont souvent sensibles
à la fois à l’orientation et à la couleur. Le rôle
exact de l’aire V4 est encore débattu, mais cette aire intervient
sans doute dans la reconnaissance des formes et semble essentielle pour la perception
des couleurs.
L’aire IT, qui tient son nom du
cortex inféro-temporal où elle est située, comprend les aires
TEO et TE. Les cellules de l’aire IT reçoivent de nombreuses connexions
de l’aire V4 et répondent à une vaste gamme de couleurs et
de formes géométriques simples. Elles joueraient un rôle important
dans la mémoire visuelle, en plus d’être une région
clé dans la reconnaissance des objets.
On a découvert dans l’aire IT des
neurones qui répondent spécifiquement à la présentation
d’images de visage. Ces enregistrements cellulaires faits chez le singe
ont été corroborés par des études d’imagerie
par résonance magnétique fonctionnelle chez l’humain. Cette
découverte n’est pas sans intérêt pour les neuropsychologues
qui connaissent depuis longtemps un syndrome rare dénommé prosopagnosie
où les patients ont de la difficulté à reconnaître
les visages malgré le fait que le reste de leur vision est normale. De
plus, on sait que ce syndrome apparaît justement à la suite de lésions
des aires extrastriées du cortex visuel.