En imagerie cérébrale,
lorsqu’on demande à quelqu’un de bouger son pouce, on observe
une activation des aires somatosensorielles et pariétales postérieures,
de l’aire 8 du cortex préfrontal et des aires motrice 4 et 6. Il
est intéressant de noter que si l’on demande au sujet uniquement
de répéter
le mouvement mentalement sans l’effectuer réellement, l’aire
6 est toujours activée, mais pas l’aire 4.
Des dommages au cortex pariétal
postérieur peuvent donner naissance à un syndrome appelé
apraxie. Selon le type d’apraxie, les patients auront par exemple de la
difficulté à faire certains gestes sur demande alors qu’ils
peuvent effectuer ce même geste spontanément. Ou encore, ils ne savent
pas faire correctement les gestes pour utiliser des objets comme un crayon ou
des ciseaux, dont ils décrivent pourtant parfaitement la fonction.
De plus, ces personnes auront d’autant plus de difficulté qu’on
leur demandera d’exécuter le geste en dehors du contexte social approprié.
Ce qui est altéré semble donc être la capacité de faire
des mouvements volontaires qui ne sont pas directement suscités ou stimulés
par l’environnement.
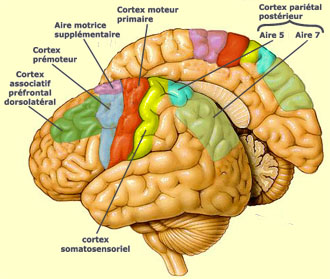
LE CORTEX MOTEUR
La région anatomique
du cerveau correspondant à l’aire 4 a reçu le nom de cortex
moteur primaire (ou M1) depuis que Penfield
a montré que des stimulations focales dans cette région évoquaient
des contractions musculaires très localisées à différents
endroits du corps. Celui-ci est représenté de façon somatotopique
sur le cortex moteur avec pondération de l’étendu des surfaces
en fonction de la précisions des mouvements pouvant être évoqués
par chaque partie du corps (voir encadré sous ce texte).
Le
cortex moteur comprend également, rostralement à l’aire 4,
l’aire 6 qui se subdivise en aire prémotrice et en aire motrice supplémentaire.
Le cortex prémoteur interviendrait dans la régulation de la posture,
en dictant au cortex moteur une position optimale pour un mouvement donné.
L’aire motrice supplémentaire, quant à elle, semble influencer
la planification et l’initiation des mouvements en fonction des expériences
passées. Le simple fait d’anticiper un mouvement déclenche
une transmission nerveuse dans la zone supplémentaire.
Outre
le cortex frontal, l’implication du cortex pariétal postérieur
dans le mouvement volontaire ne fait aucun doute. Elle se joue dans l’évaluation
du contexte dans lequel s’effectue le mouvement. Le cortex pariétal
évalue ainsi différentes données comme la position du corps
et de la cible dans l’espace grâce aux informations somatosensorielles,
proprioceptives et visuelles qu’il reçoit. Il produit ainsi des modèles
internes du mouvement à effectuer, en amont des cortex prémoteur
et moteur.
On distingue deux aires particulières dans le cortex
pariétal postérieur. L’aire 5 reçoit les informations
des aires corticales somatosensorielles 1, 2 et 3. L’aire 7, quant à
elle, intègre des signaux déjà fortement intégrés
en provenance des
aires visuelles comme MT (ou V5).
Les lobes pariétaux sont
eux-mêmes étroitement interconnectés avec les
aires préfrontales qui représentent avec eux le plus haut degré
d’intégration dans la hiérarchie du contrôle moteur.
C’est là que les décisions sur les actions à effectuer
sont prises. Les aires pariétales postérieures et préfrontales
envoient leurs axones vers l’aire 6 qui, une fois renseignée sur
le type d’action à réaliser, aide à déterminer
les caractéristiques du mouvement approprié.
Le processus à l’origine
d’une réponse motrice volontaire est aussi fabuleusement complexe
que les systèmes sensoriels qui ont fourni l’information visuelle
ou auditive ayant mené au mouvement. Les fonctions motrices ont d’ailleurs
plusieurs points en commun avec les mécanismes sensoriels, et en particulier
les sensations tactiles. Ainsi, l’aire motrice corticale située dans
la partie postérieure du lobe frontal se retrouve juste à côté
de l’aire somatosensorielle localisée, elle, dans la partie la plus
antérieure du lobe pariétal.
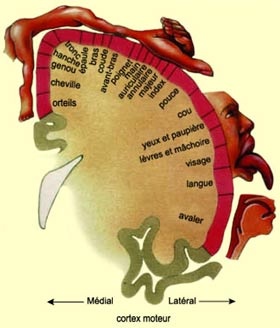
Les deux régions allongées
sont donc face à face et les fibres nerveuses qui y partent ou y arrivent
ont un même organisation somatotopique, c’est-à-dire
une cartographie qui reproduit à petite échelle l’anatomie
du corps humain. Tant pour le cortex moteur que pour le somatosensoriel, l’image
du corps que l’on peut « lire » sur le cortex est déformée
: par la précision avec laquelle est contrôlée la partie du
corps en question dans un cas, et par le degré de sa sensibilité
dans l’autre.
Bien entendu, des mouvements très fins d’autres
parties du corps peuvent être appris, tel celui du poignet, du coude et
de l’épaule du violoniste. Cela signifierait donc que la surface
corticale correspondant à ces mouvements peut s’agrandir avec l’apprentissage.
Plusieurs observations en ce sens ont d’ailleurs été faites
lors d’expérience de microstimulation corticale chez le rat. Par
exemple, la région du cortex moteur normalement impliquée dans la
commande des mouvements des vibrisses peut déclencher des mouvements du
membre antérieur quand les nerfs moteurs qui innervent les muscles du museau
sont sectionnés.
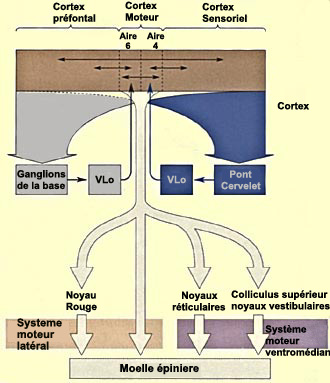
Les ganglions de la base ont un
rôle indirect dans le système moteur. En se projetant à la
fois sur le cortex moteur, prémoteur et sur l'aire motrice supplémentaire,
ils participent à une boucle dite "squeletto-motrice" qui détermine
et contrôle quels mouvements seront effectués. Leur dysfonction entraîne
soit une perte de mouvements (hypokinésie) comme c’est le cas avec
un syndrome parkinsonien, ou soit un excès de mouvements (hyperkinésie),
comme lors d’une chorée de Huntington.
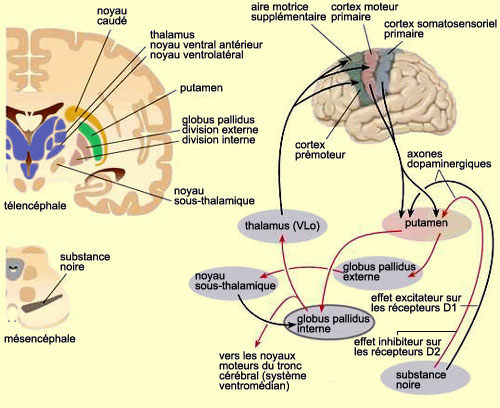
LES GANGLIONS DE LA BASE
L’ensemble
formé par le
noyau caudé (en orange) et le putamen (en vert) est appelé striatum.
Il constitue la cible majeure des afférences corticales aux ganglions de
la base. Les efférences qui vont vers le thalamus partent pour leur part
du globus pallidus. La partie du noyau ventro-latéral du thalamus qui projette
par la suite vers l’aire 6 est appelée « pars oralis »
et désignée habituellement par l’abréviation VLo.
Les
autres structures des ganglions de la base participent à des boucles internes
diverses qui modulent l’activité du circuit principal où l’information
traverse successivement les structures cérébrales suivantes: cortex
– striatum – globus pallidus – VLo – cortex (aire motrice
supplémentaire ou AMS).
On connaît la nature excitatrice ou
inhibitrice des connexions de cette boucle principale. Elles sont représentées
sur le schéma ci-bas, ainsi que l’influence excitatrice de la substance
noire et du noyau sous-thalamique sur des régions différentes de
ce circuit.
Source : Jacob L. Driesen, Ph.D.
En
considérant le sens des interactions entre les différentes structures
de cette boucle, on en arrive à comprendre globalement comment elle fonctionne.
On sait par exemple qu’au repos, les neurones du globus pallidus sont spontanément
actifs et exercent par conséquent une inhibition sur le VLo du thalamus.
Or, quand survient une activation de la boucle en provenance du cortex, les neurones
du putamen sont activés et inhibent subséquemment ceux du globus
pallidus. Ces derniers, étant tout à coup moins actifs, libèrent
les cellules du VLo de leur inhibition pallidale. L’activation du VLo qui
en résulte a pour effet de faciliter l’activité de l’AMS.
Cette boucle en est donc une de rétroaction positive susceptible
de focaliser des informations issues de larges régions corticales sur l’aire
motrice supplémentaire. On peut alors imaginer que le signal déclenchant
éventuellement l’exécution du mouvement volontaire se produit
lorsque l’activation de l’AMS atteint un certain seuil sous l’influence
de cette boucle.
Parmi les différentes activités
où le cervelet serait impliqué dans l’apprentissage, on peut
mentionner l’adaptation d’un certain nombre de réflexes comme
le réflexe vestibulo-occulaire. Il s’agit du réflexe
qui permet de maintenir la direction du regard lorsqu’on tourne la tête
et que les yeux se déplacent en sens inverse. Or ce réflexe est
modifiable avec l’apprentissage et des lésions au cervelet empêchent
cet apprentissage.
L’apprentissage d’un conditionnement est
aussi perturbé par des lésions au cervelet. Le réflexe
palpébral, qui fait fermer automatiquement notre paupière lorsqu’un
jet d’air agresse notre œil, en fournit un exemple. On peut conditionner
ce réflexe en présentant un son juste avant le jet d’air.
Après plusieurs associations de ce genre, le son seul provoque la fermeture
de la paupière. Or un cervelet endommagé supprime ce réflexe
conditionné ou empêche son apprentissage.
Le cervelet possède dans
ses circuits un système capable de mesurer le temps. Cette capacité
lui permet d’ordonner différentes fonctions qui sont sous son contrôle.
Une personne dont le cervelet est endommagé a par exemple beaucoup de difficulté
à estimer un intervalle de temps entre deux sons et de le comparer à
un intervalle témoin.
Le défaut de ce système de
mesure du temps expliquerait le calcul erroné, à partir de la
vision, de la vitesse du déplacement des objets ou des segments corporels.
Il rendrait également compte de la mauvaise coordination motrice, que ce
soit des phases d’accélération ou de freinage du mouvement.
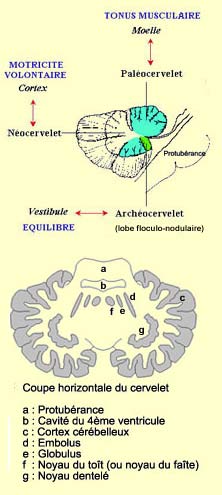
LE CERVELET
Les
pathologies du cervelet nous ont révélé depuis longtemps
que cette structure cérébrale est impliquée dans la coordination
motrice (voir encadré). Le cervelet est subdivisé en trois régions.
Chacune de ces régions est connectée à une structure cérébrale
spécifique et impliquée dans une fonction précise.
L'archéocervelet
(ou vestibulocerebellum), qui a fait son apparition pour la première
fois chez les poissons, est connecté au vestibule et est impliqué
dans l'équilibre.
Le paléocervelet(ou spinocerebellum) comprend
surtout la partie axiale du cervelet appelé vermis et se superpose
au précédent chez les amphibiens, les reptiles et les oiseaux. Il
et est connecté à la moelle et contrôle l’activité
musculaire de la posture en influençant le tonus musculaire. Le muscle
doit en effet être tendu pour jouer son rôle de maintien corporel.
Le cervelet contrôle donc à chaque instant la tension musculaire
et laisse libre les muscles chargés de faire des mouvements.
Le néocervelet(ou cerebrocerebellum)
se superpose aux précédents chez les mammifères
et est spécialement volumineux chez les primates et
surtout chez l’homme. Constitué par les hémisphères
cérébelleux, il est connecté au cortex
et contribue à la coordination des mouvements volontaires.
Il s’assure entre autres que les muscles antagonistes
freinent les muscles à l’origine du mouvement
pour atteindre parfaitement le but.
Dans le cervelet, la substance grise est
aussi organisée un peu comme dans les hémisphères cérébraux
: un cortex qui forme la matière grise à la surface et des noyaux
en profondeur qui servent de relais pour les voies efférentes qui partent
du cortex cérébelleux. Ces noyaux cérébelleux sont
au nombre de quatre de chaque côté de la ligne médiane : les
noyaux fastigiaux, aussi appelé du toit ou du faîte
(relais pour l'archéocervelet), les noyaux emboliformes
et globuleux (relais pour le paléocervelet) et les noyaux
dentelés qui sont situés au milieu de chaque hémisphère
cérébelleux (relais pour le néocervelet).
Pour pouvoir effectuer
un mouvement quelconque, il faut que l’enchaînement et la durée
des mouvements élémentaires de chaque segment corporel impliqué
dans le geste puisse être réglé de façon très
précise. Le cervelet assume, entre autres, cette fonction de réglage
temporel de nos mouvements. Il le fait grâce à un
circuit en boucle qui le relie au cortex moteur et permet la modulation du
signal qui y est envoyé en direction des motoneurones.
Chez l’humain,
le cervelet joue aussi un rôle dans l’analyse des signaux visuels
liés au mouvement. Ces signaux proviennent du déplacement d’objets
dans le
champ visuel ou encore de la vision du mouvement des segments corporels du
corps. Le cervelet interviendrait pour calculer la vitesse du déplacement
et pour ajuster en conséquence la commande motrice. Chez le patient ayant
subi une lésion au cervelet, ce calcul erroné compte pour beaucoup
dans le mauvais contrôle des mouvements.
Quant aux fonctions cognitives, des signes d’atteintes
ont été mis en évidence dans le domaine du langage, de l’attention,
de la mémoire et de l’émotion. Par exemple, chez certains
enfants autistes, les retards sur le plan cognitif ont été partiellement
imputés au développement insuffisant de certaines parties du cervelet.
Le « syndrome cérébelleux
» est l’expression employée pour désigner les manifestations
d’une atteinte au cervelet, quel que soit son origine (lésion, tumeur,
accident cérébro-vasculaire, etc.). Si par exemple, un patient souffrant
de syndrome cérébelleux veut toucher un objet, le mouvement de sa
main démarre avec du retard, puis s’accélère au-delà
de la normale, de sorte la trajectoire est déviée au-delà
de l’objet. Ce mouvement dont le freinage est trop tardif et manque d’efficacité
se termine par des oscillations du bras et de la main.
Des troubles de
la coordination se manifestent aussi dans le domaine de la posture et de l’équilibre
où la personne présente une démarche incertaine avec un élargissement
de l’écart entre les pieds lors de l’appui au sol. Si ces patients
son bousculés, les réflexes qui compensent le déséquilibre
sont exagérés et entraînent souvent des oscillations de tout
le corps. Ces personnes ne peuvent pas non plus incliner leur tronc vers l’avant
ou vers l’arrière sans perdre inévitablement l’équilibre.
Le fonctionnement de chaque niveau
hiérarchique de la commande motrice est extrêmement dépendant
de l’information sensorielle qu’il reçoit. Tellement, en fait,
que le système moteur doit plutôt être considéré
pleinement sous son aspect sensorimoteur. Au niveau de la détermination
des stratégies motrices, les informations sensorielles contribuent à
générer une image mentale du corps et de sa situation dans l’environnement.
Les décisions sur les paramètres à adopter pour la commande
motrice (durée, amplitude de contraction, etc) sont basées sur la
mémoire des informations sensorielles relatives aux mouvements précédents.
Et au niveau de l’exécution comme telle du mouvement, la rétroaction
sensorielle permet le maintient postural et aide à déterminer la
longueur et la tension des muscles avant et après chaque mouvement volontaire.
LA SÉQUENCE D'ACTIVATION DES AIRES MOTRICES
On peut dire que le mouvement
volontaire est avant tout un effort intentionnel entrepris conjointement par le
cortex moteur et de nombreux « consultants » nerveux. Cet effort est
organisé de façon hiérarchique où un premier niveau
s’occupe à définir les stratégies motrices, c’est-à-dire
les objectifs du mouvement et les comportements à mettre
en œuvre pour atteindre ces objectifs. Décider de prendre l’ascenseur
par exemple, qui implique de se rapprocher du bouton d’ascenseur pour éventuellement
appuyer dessus, sollicite le cortex préfrontal où s’élabore
la planification du mouvement. Le cortex frontal reçoit lui-même
un important contingent d’axones du cortex pariétal. Celui-ci étant
impliqué dans la perception spatiale, son analyse de la position du corps
et des différents membres dans l’espace est essentielle à
la préparation d’un mouvement. Les
ganglions de la base sont également des structures cérébrales
impliquées à ce niveau.
Dans
un second temps, les
aires motrice secondaires (APM et AMS) travaillent avec le
cervelet à spécifier la séquence précise de
contractions des différents muscles requis pour la réalisation
de l’acte moteur choisi, en l’occurrence ici soulever le bras et tendre
l’index vers le bouton d’ascenseur. Pour ce faire, il faudra cependant
transformer l’emplacement du bouton d’ascenseur dans l’environnement
extérieur en coordonnées intrinsèques permettant d’ajuster
les angles des différentes articulations participant au mouvement.
Dans
un troisième temps, le cortex
moteur primaire, le tronc cérébral et la moelle épinière
entrent en jeu pour produire la contraction de tous les muscles nécessaires
au mouvement choisi. Le cortex moteur primaire détermine la force que chaque
groupe de muscles doit exercer et transmet ensuite ces informations aux motoneurones
et aux interneurones spinaux qui génèrent le mouvement en tant que
tel ainsi que les ajustements posturaux qui l'accompagnent.
Appliqué au mouvement de lancer d’une
balle par exemple, ces trois niveaux s’intégrerait un peu de la façon
suivante. Le cortex cérébral est d’abord informé de
la position du corps dans l’espace par l’entremise de l’information
sensorielle visuelle, auditive, somatique et proprioceptive qu’il reçoit.
Le cortex effectue des échanges d’information avec les ganglions
de la base quant à l’objectif visé par le lancé (lancer
loin, haut, fort ?) et la stratégie à adopter selon entre autre
l’expérience du sujet dans ce domaine. Les aires motrices secondaire
du cortex cérébral et le cervelet prennent alors des décisions
appropriées concernant l’amplitude, la direction et la force du mouvement
à effectuer par le bras. Ils transmettent ces instructions au tronc cérébral
et à la moelle épinière cervicale qui va provoquer un mouvement
coordonné de l’épaule, du coude, du poignet et des doigts
de la main. Simultanément, les ordres donnés à la moelle
épinière thoracique et lombaire à partir du tronc cérébral
déterminent des ajustements posturaux qui vont permettre à la personne
de garder son équilibre tout en optimisant son mouvement durant le lancer
de la balle. Les motoneurones du tronc cérébral sont également
activés pour maintenir le regard du lanceur sur sa cible.