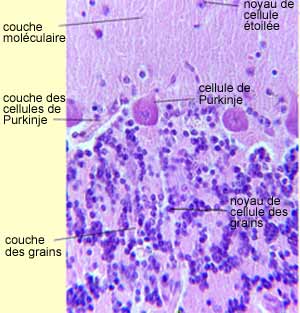
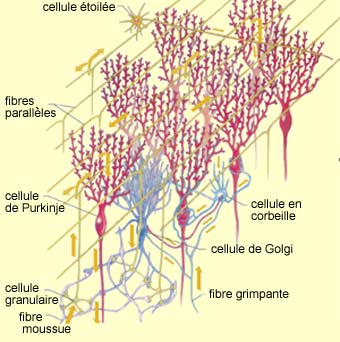
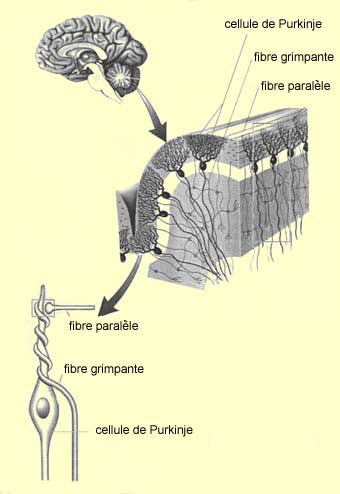
|
|
| | | |
 |
| Produire un mouvement volontaire |
| |
| | | |



Les réflexes spinaux, comme
le réflexe
rotulien ou le réflexe
de retrait d’une jambe, révèlent déjà toute
la complexité du système spinal de contrôle du mouvement.
L’activité des motoneurones alpha qui innervent les muscles est sous
la triple influence des afférences sensorielles, des interneurones spinaux
et de la voie pyramidale. |
On a pensé pendant un certain
temps qu’il existait une représentation détaillée de
chaque muscle du corps dans le cortex primaire, de sorte que l’activation
d’une cellule pyramidale particulière conduisait à l’activation
d’une seule population de motoneurones. Cette vision des choses est toutefois
remise en question par des travaux récents suggérant que les cellules
pyramidales commanderaient plutôt un groupe de muscles. Leur activation
permettrait alors de mobiliser un membre tout entier vers un objectif.
L’enregistrement de l’activité des neurones du cortex moteur
durant un mouvement montre par ailleurs une activation qui débute avant
le mouvement et se maintient durant son exécution. Cette activité
est susceptible de coder deux aspects principaux du mouvement : sa force et sa
direction. 
|

Nos
neurones miroirs préfèrent nos mouvements préférés
Huit
problèmes avec les neurones miroirs
La
cote des cellules gliales en hausse, celle des neurones miroirs
en baisse
Notre
corps à l’origine de notre compréhension du langage
Daniel
Glaser : un neuroscientifique qui explique
La
danse (comme la musique) modifie le cerveau de ceux qui en font
souvent
|
|
| LES AXONES QUI ENTRENT ET SORTENT DU CORTEX MOTEUR |
| En plus de recevoir des
afférences des aires prémotrices et motrices supplémentaires,
les neurones pyramidaux du cortex moteur primaire reçoivent de l’information
directement des aires somatosensorielles 3, 1 et 2. L’autre entrée
importante d’axones provient du thalamus, plus précisément
d’une partie du noyau ventrolatéral dénommée VLc et
relayant l’information issue du cervelet.
L’efférence
principale du cortex moteurs primaire est la voie corticospinale avec son système
latéral et son système ventromédian.
Le système latéral est
formé de deux faisceaux principaux dont le plus important est la voie
corticospinale latérale. Prenant naissance surtout dans les aires
4 et 6 du lobe frontal, désigné globalement comme le cortex moteur,
c’est la voie la plus longue et une des plus importante en terme de quantité
d’axones (un million d'axones). Les autres axones de ce faisceau sont principalement
issus des aires somatosensorielles et du lobe pariétal.
Après avoir traversé la capsule interne, le mésencéphale
et le pont, les axones issus du cortex se réunissent
au niveau du bulbe pour former un dense faisceau de fibres nerveuses.
Celui-ci a un peu la forme d’une pyramide s’étendant
à la surface ventrale du bulbe rachidien, d’où
son nom de faisceau pyramidal à ce niveau.
Par opposition, toutes les autres voies en provenances des structures
sous-corticales ou du tronc cérébral qui passent
ailleurs reçoivent souvent le nom de voies extrapyramidales.
Juste avant d’entrer
dans la moelle épinière, le faisceau pyramidal présente une
décussation. Autrement dit, les fibres en provenance de l’hémisphère
gauche du cortex vont maintenant cheminer dans la colonne latérale droite
de la moelle épinière, et vice-versa. Les terminaisons axonales
de cette voie rejoignent finalement les motoneurones et les interneurones de la
partie dorsolatérale de la corne ventrale de la moelle épinière.
Le faisceau rubrospinal est le second faisceau du système
latéral. Il provient des neurones du noyau rouge situé dans le mésencéphale.
Ce noyau reçoit des informations du cortex frontal, une région qui
contribue déjà massivement à la voie corticospinale. Au
cours de l’évolution des primates, on observe justement une diminution
de cette voie indirecte au profit d’une prise en charge corticospinale de
plus en plus importante du contrôle moteur. | 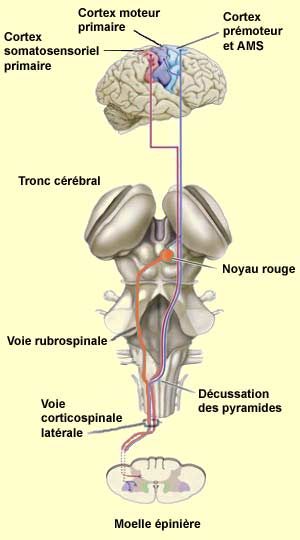
|
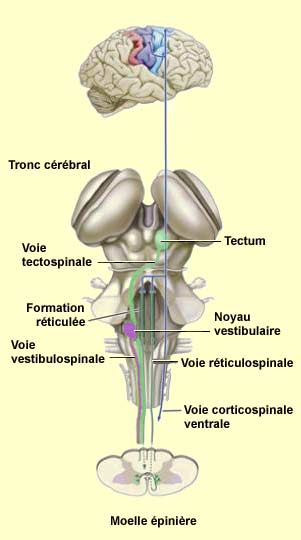 | L’autre
grande voie descendante, le système ventromédian, est constitué
de quatre faisceaux qui prennent leur origine dans diverses régions du
tronc cérébral et contribuent essentiellement au contrôle
postural et à certains mouvements de type réflexe. Les neurones
à l’origine de ces faisceaux reçoivent des informations sensorielles
en rapport avec le sens de l’équilibre, la position du corps et l’environnement
visuel.
Ainsi, le faisceau vestibulospinal est issu
des noyaux vestibulaires qui reçoivent de l’information de l’oreille
interne. Il contribue à maintenir la tête en position correcte par
rapport aux épaules, ce qui est essentiel pour conserver une stabilité
du regard lorsque le corps se déplace.
Le faisceau tectospinal
prend son origine du collicule supérieur du mésencéphale
(tectum) qui reçoit des informations directes de la
rétine, mais aussi des informations somatosensorielles et auditives.
À partir de la représentation de l’environnement que forme
le collicule supérieur, cette voie contribue à l’orientation
du regard.
Les faisceaux réticulospinaux d’origine
pontique et bulbaire proviennent quant à
eux de la formation réticulée du tronc cérébral. Recevant
des informations de toute origine, la formation réticulée s’étend
sur toute la longueur du tronc cérébral, du pont au bulbe rachidien.
L’action de ces voies contribue au maintient de la posture, les axones d’origine
pontique favorisant les réflexes antigravitaires spinaux, et les axones
d’origine bulbaire libérant au contraire les muscles qui s’opposent
à la gravité, facilitant ainsi d’autres mouvements. |
Dans l’aire F5 du cortex prémoteur
ventral du singe, on a découvert, vers le milieu des années 1990,
que certains neurones émettaient des potentiels d’action non seulement
lorsque le singe faisait un mouvement de la main ou de la bouche, mais aussi lorsqu’il
regardait simplement un autre animal ou un humain faire le même geste. On
appela ces neurones des « neurones miroirs » parce
que l’action observée semble reflétée, comme dans un
miroir, dans la représentation motrice de la même action chez l’observateur.
Outre les neurones miroirs qui s’activent lorsque nous voyons se réaliser
la même action que celle pour laquelle ils sont impliqués quand nous
la faisons, un autre type de neurones dits «canoniques» s’activent
quant à eux à la simple vue d’un objet saisissable par le
mouvement de préhension de la main codé par ce neurone. Comme si
cerveau anticipait une interaction possible avec cet objet et se préparait
en conséquence.
Ces deux types de neurone ont cependant en commun
d'être activés par une action, qu'elle soit effectuée, vue
ou anticipée. Parce que nous pouvons prévoir les conséquences
de nos propres actions, certains ont avancé que les neurones miroirs pourraient
être le substrat neuronal de notre capacité à comprendre également
la signification d’une action faite par autrui.
Or cette compréhension
des actions de l’autre est à la base des relations sociales et particulièrement
de la communication interindividuelle. Cette découverte revêt donc
un caractère extrêmement intéressant pour expliquer comment
on peut se
représenter l'état d’esprit et les intentions des autres.
Enfin, le fait que l’aire F5 chez le singe est considérée
comme l’homologue de l’aire de Broca chez l’humain suggère
aussi une implication des neurones miroirs dans la communication humaine. | |
|