|
|
| Un livre accompagne le site depuis octobre 2024. Découvrez-le ici > | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|


|
|
Des souvenirs toujours reconstruits Intégration et conscience : jamais deux sans trois ! Norman Doidge et la plasticité cérébrale
Faire
des liens grâce à l’école d’été
sur le raisonnement
| |
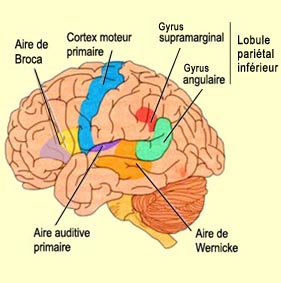
V.S. Ramachandran suggère par exemple qu’un nouvel ensemble de structures cérébrales aurait évolué durant l’hominisation pour pouvoir transformer les outputs des aires sensorielles primaires en ce qu’il appelle des « métareprésentations ». Autrement dit, au lieu de produire de simples représentations sensorielles, le cerveau s’est mis à créer « des représentations de représentations » qui vont rendre possible la pensée symbolique. Et c’est sous cette forme bonifiée que l’information sensorielle deviendrait plus aisément manipulable, notamment pour le langage.
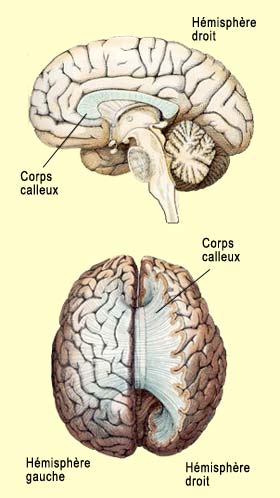
Pour Ramachandran, l’interaction de l’aire de Wernicke du lobule pariétal inférieur, en particulier le droit, et du cortex cingulaire antérieur, est fondamentale pour générer des métareprésentations à partir des représentations sensorielles, donnant ainsi lieu aux qualia et au sentiment d’un « soi » qui ressent ces qualia. Mais si l’hémisphère droit, à cause de son implication dans l’image globale du corps, est très important pour générer ce sentiment d’être « soi », l’hémisphère gauche, celui qui est spécialisé dans le langage chez la grande majorité des gens, semble être à sa façon tout aussi essentiel à ce que nous appelons la conscience de soi.
Michael Gazzaniga a mis en évidence plusieurs situations où les deux hémisphères cérébraux des patients à cerveau divisé entrent en conflit (voir la capsule expérience ci-bas pour des exemples). De façon générale, si l’on envoie un stimulus visuel à l’hémisphère droit via l’oeil gauche, le sujet affirme n’avoir rien vu mais peut identifier l'objet parmi d'autres si on lui donne à palper avec la main gauche (sous contrôle de ce même hémisphère droit), sans toutefois pouvoir le nommer. Tout se passe comme s’il y avait deux individus dans la même personne; un premier qui dépend de l’hémisphère gauche et peut prend la parole; et un second qui dépend de l’hémisphère droit, qui est privé de parole, mais qui fait la preuve qu’il perçoit et agit en conséquence si on l’interroge correctement.
Cet interprète nous raconterait constamment une histoire cohérente construite à partir de nos actions, nos émotions et nos pensées. Ce serait en quelque sorte la « colle » permettant de mettre ensemble les différents éléments de notre histoire personnelle et de nous donner l’impression d’être un agent rationnel. En fait, pour Gazzaniga, il est même possible que l’hémisphère droit ait aussi son propre interprète. Même s’il ne parle pas, cet observateur de l’hémisphère droit serait capable de mieux comprendre l’ironie, les blagues et d’autres stratégies émotionnelles. Chacun de ces deux interprètes contrôlerait également les mouvements volontaires des membres du côté opposé du corps. Est-ce qu’un patient au cerveau divisé possède alors deux consciences ? Et chez la personne normale, y’aurait-il également ces deux consciences qui seraient cependant intimement reliées pour nous donner cette impression de n’être qu’une seule et même personne ? La présence d’un « interprète », d’une espèce d’observateur dans le cerveau, ne fait en tout cas pas consensus et certains, comme Daniel Dennett, l’ont fortement critiqué. Chose certaine, les expériences avec les cerveaux divisés ont fait couler beaucoup d’encre et n’ont pas fini d’en faire couler…
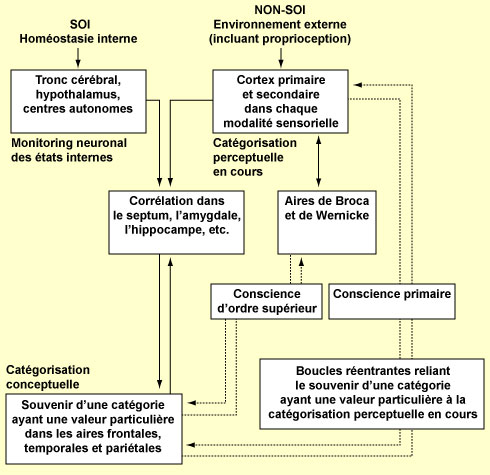
Ceci dit, si certains modèles neurobiologiques de la conscience ciblent des structures particulières du cerveau comme le lobule pariétal inférieur ou mettent plutôt l’emphase sur un hémisphère en entier, d’autres en contrepartie affirment carrément que la conscience n’est le fait d’aucune structure précise du cerveau. Rappelons que l’étude des bases neurobiologiques de la conscience a souvent été abordée comme celle de toute autre fonction, c’est-à-dire en analysant la contribution de différentes régions cérébrales ou de différents groupes de neurones. Mais plusieurs ont critiqué cette approche conventionnelle qui, selon eux, n’est pas pertinente pour étudier une fonction aussi globale que la conscience. Ils rappellent d’une part que des dommages à pratiquement n’importe quelle structure cérébrale a des effets sur la conscience. Et d’autre part, que des lésions localisées affectent la conscience, mais l’anéantit rarement complètement. Un autre fait, souligné par Gerald M. Edelman et Giulio Tononi, est que les circuits de notre cerveau sont très redondants (« massively degenerate », en anglais) et que plusieurs circuits peuvent accomplir la même fonction. Edelman et Tononi font donc partie de ceux qui appuient l’idée que le cerveau en entier aurait son mot à dire dans les processus conscients. Leur modèle vise à expliquer deux caractéristiques de la conscience qui leur semblent fondamentales : le fait que chaque état de conscience est un tout indivisible, et qu’en même temps chaque personne peut choisir à tout moment entre d’innombrables états conscients. En d’autres termes, leur modèle veut rendre compte de l’unité de la conscience et de sa complexité. Comme la plupart des neurobiologistes qui tentent de modéliser la conscience, Edelman et Tononi se voient eux aussi contraints de distinguer au moins deux grandes formes de conscience, la conscience primaire et la conscience d’ordre supérieur. La conscience primaire que nous partageons avec plusieurs autres animaux est celle qui permet de prendre conscience de la situation dans laquelle nous nous trouvons. Elle fait appel à une mémoire à court terme qui rend possible le « présent remémoré », pour employer l’expression d’Edelman. Lorsqu’un stimulus est reçu par l'organisme, il y a formation de cartes perceptuelles constituées d’assemblées de neurones dont les connexions réciproques sont renforcées. Il en résulte un système de « cartes neuronales », chacune responsable de nos différentes possibilités perceptuelles, qui sont issues d’un processus sélectif appelé « Darwinisme neuronal » par Edelman (voir le premier encadré). Quand le cerveau reçoit une nouvelle stimulation, plusieurs de ces cartes vont être activées et vont s’envoyer des signaux mutuels. La conscience primaire naîtrait de l’interaction de différentes assemblées neuronales codant pour différentes propriétés d’un objet. Interaction qui se ferait surtout par l’entremise de connexions réciproques formant des boucles qui peuvent unir des groupes de neurones parfois très éloignés dans le cerveau. La conscience primaire dépendrait donc à chaque instant de l’activité parallèle et récursive à l’intérieur et entre des régions du thalamus et du cortex. Même s’ils ne cherchent pas à faire une association trop étroite entre certaines structures cérébrales et les états conscients, Edelman et Tononi reconnaissent tout de même l’importance indéniable des boucles thalamo-corticlales dans l’émergence de la conscience, bien qu’ils accordent moins d’importance à la synchronisation neuronale en tant que telle qu’à la signification fonctionnelle de ces boucles. La conscience d’ordre supérieur, qui émerge durant l’hominisation, dépendrait elle aussi de ces «boucles réentrantes» entre des assemblées de neurones. Mais des boucles qui se situeraient à plus grande échelle dans le cerveau, en particulier entre les aires corticales associées au langage et celles associées aux concepts abstraits. L’explosion des capacités sémantiques qui s’ensuit permettrait alors l’émergence du concept de soi, et donc la considération de la conscience primaire à la lumière du passé et du futur. Autrement dit, l’émergence d’une conscience d’avoir conscience, ce qui est la définition de la conscience d’ordre supérieur. Concrètement, tout cela serait rendu possible par la constitution d’un «noyau dynamique», c’est-à-dire une vaste assemblée de neurones en réarrangement constant, mais qui maintient une continuité et est aussi capable d’intégration à travers la complexité (les deux caractéristiques principales de la conscience dont ce modèle veut rendre compte). En d’autres termes, le noyau dynamique permettrait de rendre compte du fait qu’à tout moment, il n’y a qu’un seul objet conscient, mais que la conscience peut passer très rapidement d’un objet à l’autre. Tout comme Llinás ou Freeman, Edelman et Tononi insistent donc sur l’aspect dynamique de leur noyau qui réfère ici non pas à une structure anatomique stable mais bien à un pattern d’activité éphémère impliquant les neurones de différentes régions du cortex à un moment donné. Et comme à tout moment, le noyau dynamique peut impliquer des régions du cerveau complètement différentes, cette hypothèse évite donc de devoir désigner une région cérébrale particulière, ou un type de neurone particulier, ou encore une fréquence d’activité neuronale particulière qui serait associée à nos états de conscience. À la place, ce modèle propose qu’une activité neuronale participe à un état conscient si elle se trouve incluse, au moment où cet état est conscient, dans le noyau dynamique. Cette hypothèse a donc beaucoup de chose en commun avec les théories de l’espace de travail global, bien qu’elle s’en distingue par le fait qu’un contenu n’est pas rendu conscient simplement parce qu’il est rendu disponible au reste du système. Dans l’hypothèse du noyau dynamique, il n’y a pas l’équivalence entre accessibilité et conscience que l'on retrouve dans les théories de l’espace de travail global. Bien que les neurones de ce système dynamique interagissent principalement entre eux, ils influencent et sont influencés par d’autres structures cérébrales. En effet, même si le modèle de la conscience d’Edelman et Tononi, contrairement à d’autres, ne cherche pas à mettre certaines régions du cerveau «en vedette», plusieurs structures cérébrales y sont mentionnées comme jouant un rôle important.
C’est le cas bien entendu du thalamus et du cortex qui participent à la formation des boucles réentrantes. Mais c’est aussi le cas des noyaux du tronc cérébral dont les axones remontent dans de vastes régions du cerveau, et qui relâchent de manière diffuse des neurotransmetteurs comme la noradrénaline, la sérotonine, l’acétylcholine ou la dopamine. Le seuil d’activation des différentes assemblées de neurones du cortex sera alors déterminé par ces systèmes de neuromodulation diffus (Edelman parle en anglais de « diffuse ascending value systems”). Certains neurones de ces noyaux neuromodulateurs vont par exemple être en activité tonique quand l’animal est éveillé et l’arrêt de cette activité va contribuer à plonger l’animal dans le sommeil. D’autres vont produire une activité phasique quand quelque chose de nouveau ou d’important pour l’animal apparaît dans l’environnement. Par exemple, les neurones du locus coeruleus relâchent dans la majorité des structures cérébrales une bouffée de neuromodulateurs, dans ce cas-ci de la noradrénaline, dès qu’un animal entre dans un nouvel environnement ou que quelque chose d’inusité survient. Ces neuromodulateurs peuvent donc modifier l’activité de vastes populations de neurones. Ils peuvent également changer la probabilité que des synapses se renforcent ou s’affaiblissent en réponse à une activité neuronale donnée. Ces systèmes de neuromodulateurs sont donc extrêmement bien conçus pour amplifier un type d’activité cérébrale au dépens d’un autre. Enfin, la capacité de choisir entre de nombreux inputs ceux qui ont une certaine importance relativement à l’histoire individuelle d’une personne constitue avantage adaptatif certain. L’hippocampe, qui a co-évolué avec le cortex et interagit avec lui pour produire ce que l’on appelle une mémoire épisodique, joue probablement un rôle clé dans ce processus.
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

| |
|
|
|
|
|
|
|
|