Une fibre musculaire est le résultat
de la fusion de plusieurs cellules durant le développement. Ce phénomène,
que l’on appelle un syncytium, explique pourquoi une seule fibre
musculaire peut contenir plusieurs centaines de noyaux. Ceux-ci sont situés
à la périphérie du cytoplasme, juste sous la membrane cellulaire
(sarcolemme).
Plus un organe nécessite
un contrôle moteur fin, plus la taille de l’unité motrice est
petite. Par exemple, un motoneurone peut déclencher la contraction de moins
de 10 fibres musculaires dans les muscles oculaires. Dans le larynx, c’est
à peine 2-3 fibres qui sont contrôlées par un motoneurone.
À l’opposé, les unités motrices du muscle du mollet
(gastrocnemius) comprennent de 1000 à 2000 fibres musculaires réparties
dans tout le muscle.
Par ailleurs, bien que la réponse d’une
unité motrice soit de type tout ou rien, la force de la réponse
du muscle entier est, pour sa part, proportionnelle au nombre d’unités
motrices activées.
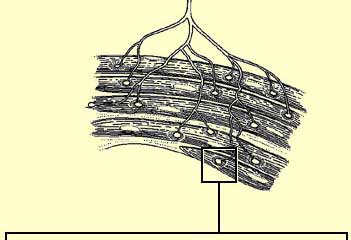
LA JONCTION NEUROMUSCULAIRE ET LA CONTRACTION DES MUSCLES
Les
axones des motoneurones,
lorsqu’ils arrivent à proximité d’un muscle pour l’innerver,
se divisent en plusieurs branches qui vont chacune établir une jonction
neuromusculaire avec une fibre musculaire. Le potentiel d’action en
provenance d’un motoneurone va donc provoquer la contraction musculaire
d’un ensemble de fibres musculaires formant ce que l’on appelle une
"unité motrice".
L'unité motrice est donc le
plus petit élément contractile que le système nerveux peut
mettre en jeu. Le corollaire étant qu’une fibre musculaire n'est
toujours innervée que par un seul motoneurone.
Quand un potentiel
d’action arrive à une jonction neuromusculaire, il provoque le relâchement
d’acétylcholine. Celle-ci se fixe sur les
récepteurs nicotiniques situé sur la "plaque motrice",
région spécialisée de la membrane post-synaptique de la fibre
musculaire où se trouve concentrés les récepteurs. Cette
fixation provoque l’ouverture de des récepteurs-canaux nicotiniques
et l’entrée d’ions sodium dans le muscle.
Cette entrée de sodium, si elle est suffisante
pour faire passer le potentiel de repos de la fibre musculaire de -95 milivolts
à environ -50 milivolts, provoque un potentiel d’action musculaire
qui se répand dans toute la fibre. Celui-ci voyage d’abord à
la surface du sarcolemme, cette membrane excitable qui entourent les différentes
structures cylindriques contractiles dénommées myofibrilles. Or,
pour atteindre les myofibrilles, dont certaines sont situées en profondeur
dans la fibre musculaire, le potentiel d’action musculaire suit le réseau
de tubule T (pour transverse) qui partent du sarcolemme et s’enfoncent
au cœur de la fibre.
Le potentiel d’action musculaire atteint
ainsi une structure clé dans la cascade de réactions menant à
la contraction musculaire : le réticulum endoplasmique, qui stocke
les ions calciums nécessaires à la contraction musculaire. Or il
existe un couplage physique direct entre une protéine sensible au potentiel
de membrane des tubules T et un canal calcique du réticulum endoplasmique,
de sorte que l’arrivée du potentiel d’action musculaire provoque
l’expulsion de calcium du réticulum endoplasmique, le rendant ainsi
disponible pour la
suite de la cascade biochimique impliquant les protéines contractiles des
myofibrilles.
L’excitabilité membranaire
est une caractéristique que partagent les neurones et les fibres musculaires
qui peuvent produire des potentiels
d’action. Cette caractéristique ne leur est cependant
pas exclusive puisqu’elle se retrouve entre autres dans les cellules glandulaires,
les ovules fertilisés et certaines cellules végétales.
Plusieurs toxines peuvent
affecter la jonction neuromusculaire et ses récepteurs nicotiniques. Certaines,
comme la toxine botulinique, agissent au niveau présynaptique en empêchant
la libération d’acétylcholine et produisant une faiblesse
ou une paralysie musculaire.
Mais d’autres agissent directement
sur le récepteur nicotinique en occupant le site de fixation de l’acétylcholine,
mais sans provoquer l’ouverture du canal. L’acétylcholine étant
libérée dans la fente synaptique mais ne pouvant se fixer aux récepteurs,
la contraction musculaire n’a pas lieu. C’est de cette façon
qu’agit le curare dont les indiens d’Amazonie enduisent leur flèche
: le poison tue en paralysant les muscles du diaphragme. C’est le même
mécanisme qui est à l’œuvre avec la bungarotoxine, un
venin de serpent.
Certaines substances vont plutôt se loger dans
le pore central du récepteur nicotinique, empêchant ainsi le passage
des ions. C’est le cas de la procaïne, la lidocaïne ou la benzocaïne,
des molécules utilisées pour l’anesthésie locale. Ou
encore de la tétrodotoxine contenue dans le foie de certains poissons et
qui, lorsqu’elle est ingérée, peut provoquer la mort en quelques
heures.
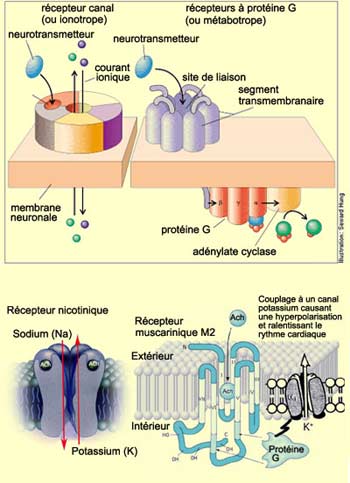
Le récepteur
nicotinique est donc un récepteur canal (ou ionotrope),
c’est-à-dire que c’est la même protéine qui forme
le canal transmembranaire et qui fixe l’acétylcholine ou l’un
de ses agonistes
comme la nicotine.
Leur fixation provoque alors une dépolarisation dans la cellule post-synaptique
en y laissant entrer beaucoup de sodium et sortir un peu de potassium.
On
distingue deux types de récepteurs nicotiniques: le N1 qui se trouve dans
le système nerveux végétatif et le N2 qui est plutôt
au niveau des jonctions neuromusculaires. Ces récepteurs font partie d’une
grande famille de récepteurs canaux qui comprend ceux de la glycine, du
glutamate
(NMDA et AMPA) ou encore du GABA (récepteurs
GABA-A).
Outre les récepteurs nicotiniques,
il existe une autre famille de récepteurs à l’acétylcholine,
les récepteurs muscariniques. Ceux-ci font partie de l’autre grande
classe de récepteur appelés récepteurs à protéine
G (ou métabotrope).
Ces récepteurs, dont font partie par exemple les récepteurs à
la dopamine, sont totalement distincts du canal ionique. Ils exercent leurs effets
sur les canaux via une protéine située du côté cytoplasmique
appelée protéine G parce qu’elle lie le GTP. Le récepteur
activé par la fixation du neurotransmetteur active donc cette protéine
G qui module directement ou indirectement (via un second messager) l'ouverture
de canaux ioniques indépendants. Son action est par conséquent moins
rapide que pour les récepteurs nicotiniques où tout est centralisé
sur le même complexe protéique.
Par ailleurs,
alors que les récepteurs nicotiniques sont formés de cinq peptides
distincts, les sept domaines transmembranaires des récepteurs muscariniques
proviennent de la même protéine qui serpente à travers la
membrane.
Encore ici, il existe au moins cinq types de récepteurs
mucariniques qui tous sont activés par la muscarine, une molécule
produite par un champignon. M1 et M3 activent par exemple la phospholipase C,
un second messager qui amène une dépolarisation par l’ouverture
de canaux au calcium et diminution du flux de potassium. M1 se retrouve dans le
cortex et les noyaux gris centraux tandis que M3 est dans le cervelet. Les deux
récepteurs interviennent également dans les sécrétions
glandulaires exocrines.
Un mode d’action différent est
associé au type M2 qui est lié à une protéine G qui
inhibe l’adényl cyclase. La baisse d’activité de cette
enzyme diminue la quantité du second messager AMP cyclique permettant l’ouverture
des canaux potassium et hyperpolarisation de la cellule. M2 se trouve également
dans le
système nerveux central (cervelet, noyaux gris centraux et tronc cérébral)
mais aussi au niveau du cœur.