|
|
| Un livre accompagne le site depuis octobre 2024. Découvrez-le ici > | |
|
|
|
|
|
||||
|
|
||||||
|
|
|
|

| |
| |
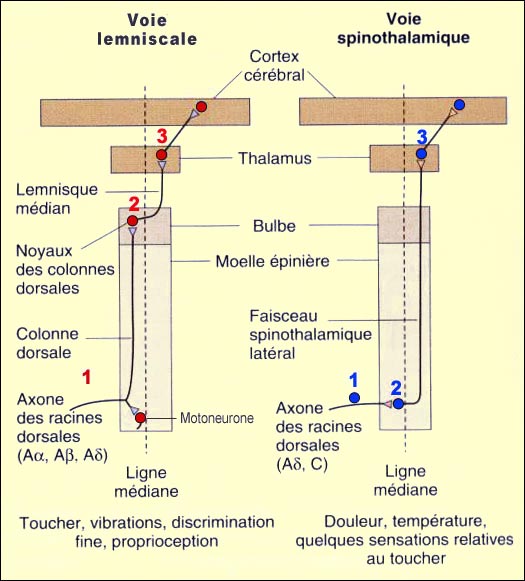
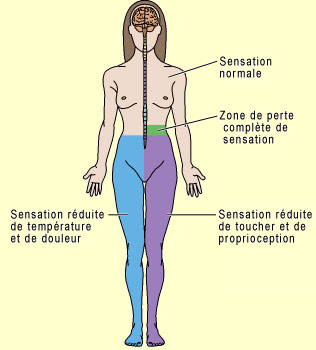
Quelles sont les voies qu'emprunte l'influx douloureux pour assurer sa fonction protectrice ? Comment l'information se rend-elle au cerveau pour nous indiquer quelle région de notre corps est endolorie ? Ces voies sont nombreuses, redondantes et complexes comme on pourrait s'y attendre pour une fonction aussi essentielle que la douleur. On doit d'abord distinguer les voies de la douleur (on dit aussi les voies nociceptives) des voies de la température non douloureuse, du toucher et de la proprioception. Deux routes différentes vont être empruntées par ces différents systèmes sensoriels. Ces routes partent toutes d'une région du corps et aboutissent au cortex somatosensoriel. Elles comptent également chacune trois neurones qui se relaient pour passer l'influx nerveux. Là où elles se distinguent cependant, c'est par l'endroit où elles vont croiser la ligne médiane dans la moelle épinière. Il faut se rappeler que dans le corps humain, tant le contrôle moteur que les afférences sensorielles sont croisés. Cela veut dire que les voies nerveuses en provenance du côté gauche du corps doivent aboutir dans l'hémisphère droit du cerveau, et vice-versa. Il faut donc que ces voies croisent la ligne médiane (on dit aussi décussent) à un moment donné. Suivons donc le cheminement d'une afférence quelconque en direction de la moelle épinière, qu'elle soit sensorielle, nociceptive, thermique ou proprioceptive. En effet, peu importe la modalité sensorielle, le corps cellulaire du premier des trois neurones de la chaîne est toujours situé dans un ganglion spinal (ou ganglion rachidien). Ceux-ci forment une chaîne de part et d'autre de la moelle épinière. On dit que ces neurones ont une forme en " T " puisqu'un court prolongement émanant du corps cellulaire se sépare rapidement en deux branches qui partent en directions opposées : l'une vers la région du corps innervée par ce nerf rachidien, et l'autre qui entre immédiatement dans la racine dorsale de moelle épinière (une zone de la moelle épinière essentiellement sensorielle, la racine ventrale étant pour sa part une zone motrice). C'est à partir de là que nos deux voies vont se distinguer.
D'après Neurosciences, M.F.Bear, B.W. Connors, M.A.Paradiso, 2001.
Tant pour pour les voies tactiles et proprioceptives que pour les voies nociceptives et thermiques non douloureuses, ce troisième neurone envoie son axone vers le cortex somatosensoriel, une région du cerveau rendant possible la localisation corporelle précise du stimulus d'origine.
|
| ||||||||||||||||||||||||||||||||||||
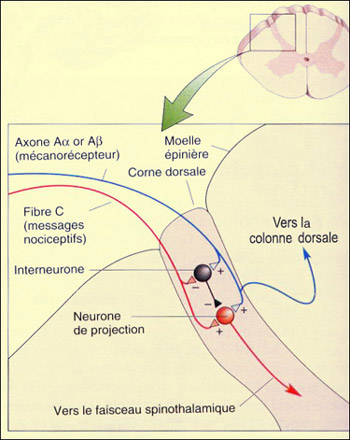
La perception douloureuse ne découle pas seulement de l'activation des voies nociceptives ascendantes mais d'un véritable dialogue de celles-ci avec les différentes voies descendantes de contrôle de cette douleur. Ce contrôle, on l'associe souvent à un système de filtre ou à des portes dont la fermeture est commandée par le cortex, le mésencéphale ou le bulbe rachidien. Mais la toute première porte, l'influx nociceptif la trouve dès son entrée dans la corne dorsale de la moelle épinière. Ce premier relais des voies ascendantes n'est donc pas seulement qu'une zone de passage de l'influx nociceptif, mais un premier lieu de filtrage de cet influx. On parle de contrôles segmentaires d'origine périphérique non douloureuse pour désigner ce premier niveau d'intégration. L'épithète " segmentaire " rappelle qu'il s'agit d'un processus qui se produit à chacun des segments de la moelle correspondant à chaque vertèbre. Ce contrôle segmentaire résulte de l'interaction entre les afférences nociceptives (A-delta et C) et non nociceptives (A-alpha et A-bêta). Cette interaction, elle a été modélisée dans un article d'abord publié en 1962, puis étoffé en 1965, que plusieurs considèrent comme le plus important à avoir jamais été écrit dans le domaine de la douleur. Leurs auteurs, le Canadien Ronald Melzack et l'anglais Patrick Wall, y proposaient le premier modèle de contrôle endogène de la douleur, la fameuse théorie du portillon (" gate control theory of pain ", en anglais). Cette théorie s'appuie sur une connectivité particulière impliquant non seulement les afférences nociceptives et celles du toucher léger comme on l'a mentionnée, mais également des interneurones inhibiteurs qui sont l'élément clé de leur explication.
C'est ce câblage particulier qui forme la porte virtuelle dont l'ouverture et la fermeture vont moduler le passage de la douleur. En effet, en situation normale, les interneurones produisent spontanément des potentiels d'action à un rythme qui leur est propre. L'activation des fibres nociceptives par un stimulus douloureux va donc d'une part stimuler le neurone non spécifique qui projette vers la voie spinothalamique (on dit aussi " neurone de projection "). Mais elle va aussi inhiber l'activité spontanée des interneurones inhibiteurs, dépolarisant ainsi le neurone de projection et augmentant d'autant plus la probabilité qu'il déclenche des potentiels d'action. Par ailleurs, l'une des premières choses que l'on fait lorsqu'on se fait mal, c'est de frotter vigoureusement l'endroit meurtri sur notre corps. Cette réaction tout à fait naturelle réduit la sensation douloureuse en " fermant " le portillon. Comment ? L'animation ci-dessous montre que si les fibres du toucher qui sont activées par le frottement excitent le neurone de projection, elle font aussi de nombreuses connexions excitatrices sur les interneurones inhibiteurs, ce qui, lorsque la stimulation tactile est soutenue, produit une forte hyperpolarisation sur le neurone de projection, diminuant ainsi de beaucoup ses probabilités d'émettre des influx nerveux.
On voit donc comment c'est le taux relatif de potentiels d'action dans les fibres nociceptives et non nociceptives qui détermine le degré d'ouverture de la " porte " au niveau de la moelle épinière, et donc le niveau de douleur qui est transmis. Par ailleurs, des projections d'origine centrale peuvent également activer ces interneurones inhibiteurs de la moelle et fermer davantage le portillon au niveau segmentaire. Même si les données recueillies depuis 1965 ont amené des modifications à la proposition originale de Melzack et Wall, l'idée d'une modulation de la perception douloureuse dès son entrée dans la moelle épinière demeure fondamentale dans le domaine du traitement de la douleur. Elle est par exemple à l'origine d'applications cliniques comme la neurostimulation transcutanée (" transcutaneous electrical nerve stimulation (TENS) ", en anglais) où l'on produit une analgésie locale en stimulant électriquement les fibres non nociceptives de la peau.
|
| |
|
|
|
|
|
|
|
|