|
|
| Un livre accompagne le site depuis octobre 2024. Découvrez-le ici > | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|


|
|
Quel chemin trace le désir ? La piste de l’ocytocine
| |
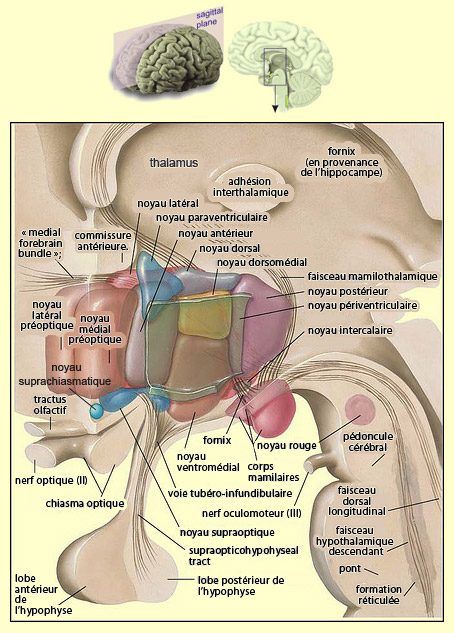
L’hypothalamus est une structure complexe du diencéphale qui permet de maintenir les fonctions vitales de l’organisme à l’intérieur de certaines limites physiologiques (ce qu’on appelle l’homéostasie). Il est intimement lié à l’hypophyse par l’entremise de ses neurohormones qui modulent les sécrétions hormonales hypophysaires et leurs multiples effets sur le corps. L’hypothalamus est également connecté à de nombreuses autres régions du cerveau afin d’assurer ce rôle de centre d’intégration des signaux pour la survie générale de l’organisme. Bien que la plupart de ces connexions soient bidirectionnelles, il est pratique de distinguer les faisceaux de fibres hypothalamiques afférentes et efférentes.
Du côté des fibres afférentes, celles qui arrivent par exemple dans les corps mamillaires représentent une voie ascendante importante pour l’hypothalamus. Elles proviennent entre autres du tegmentum du mésencéphale et de la moelle épinière. Le faisceau corticohypothalamique provient pour sa part, comme son nom l’indique, de diverses régions du cortex et se projette dans plusieurs noyaux hypothalamiques. On connaît plusieurs des routes spécifiques qui forment cette « autoroute » : celle qui part de la région corticale 6 pour aboutir dans les noyaux postérieurs et latéraux de l’hypothalamus; celle du cortex préfrontal qui se projette directement au noyau supraoptique, et indirectement en passant par le thalamus; celle qui part de la partie postérieure du cortex orbital olfactif pour se rendre jusqu’aux noyaux paraventriculaire et ventromédian; etc. Il ne fait pas de doute que c’est par l’entremise de ces voies que nous pouvons consciemment influencer notre activité viscérale, par exemple avoir mal au ventre à cause de l’anxiété ou avoir des réponses physiologiques d’excitation sexuelle par de simples pensées érotiques. Cela dit, il est certain aussi que l’hypothalamus n’est pas entièrement sous contrôle cortical, comme en fait foi notre incapacité à augmenter ou diminuer notre pression sanguine à volonté, par exemple. L’hypothalamus reçoit évidemment plusieurs fibres afférentes du thalamus situé juste au-dessus de lui. Ces fibres thalamohypothalamiques comprennent celles qui partent des noyaux antérieurs du thalamus et qui se terminent dans les corps mamillaires, ainsi que celles qui viennent surtout de la région dorsomédiale du thalamus et aboutissent dans le noyau antérieur de l’hypothalamus. L’hippocampe et l’amygdale envoient aussi des axones dans l’hypothalamus, le premier par un long faisceau recourbé appelé fornix, et la seconde par un faisceau semblable appelé la stria terminalis. La cible du fornix est constituée des corps mamillaires, tandis que celle de la stria terminalis comprend les noyaux préoptique, antérieur et ventromédian. Le tout fait partie d’une composante importante du système limbique appelé circuit de Papez. Le « medial forebrain bundle » (ou MFB) est un faisceau important de fibres nerveuses qui traverse le noyau hypothalamique latéral en se prolongeant tant rostralement que caudalement. Pour l’hypothalamus, il contient à la fois des fibres afférentes et efférentes qui le mettent en relation autant avec certaines aires corticales qu’avec le tronc cérébral.
Du côté des sorties, l’hypothalamus dispose de deux moyens pour influencer d’autres parties du cerveau et du corps : les projections nerveuses, bien sûr, mais aussi les sécrétions neuroendocrines vers l’hypophyse. Parmi les voies nerveuses, on retrouve le faisceau thalamohypothalamique déjà décrit dans les afférences. Celui-ci est en effet bidirectionnel et possède en son sein des fibres nerveuses qui vont de la région mamillaire de l’hypothalamus au noyau thalamique antérieur. De là, d’autres neurones prennent le relais et projettent leur axone au cortex cingulaire dont l’activité peut ainsi être influencée par l’hypothalamus. D’autres fibres nerveuses quittent les corps mamillaires de l’hypothalamus pour descendre dans les noyaux de la formation réticulée du tronc cérébral. Un autre faisceau important, celui des fibres périventriculaires, est constitué de fibres nerveuses qui partent notamment des noyaux postérieur et supraoptique de l’hypothalamus. Bien qu’il y ait une petite composante ascendante vers des noyaux thalamiques, la plupart de ces fibres descendent vers différents noyaux du tronc cérébral responsables entre autres de la respiration et de la pression sanguine, et même plus bas dans la moelle épinière. Les relations neurosécrétoires que l’hypothalamus entretient avec l’hypophyse sont de deux types. D’une part, la voie hypothalamo-hypophysaire, où des neurones des noyaux paraventriculaires et supraoptiques projettent leur axone dans le lobe postérieur de l’hypophyse. Ceux-ci y déversent deux neurohormones : l’ocytocine et la vasopressine. La première est sécrétée lors de l’accouchement et participe au sentiment amoureux ainsi qu’au lien social, et la seconde est impliquée dans le contrôle de la réabsorption de l’eau par les reins. D’autre part, la région périventriculaire de l’hypothalamus, par l’entremise de ses neurones dits parvocellulaires, est la source d’un second type de sortie neurosécrétoire de l’hypothalamus, cette fois-ci en direction du lobe antérieur de l’hypophyse. Ces neurones projettent leur axone dans le système porte hypothalamo-hypophysaire, un réseau de capillaires sanguins grâce auquel les facteurs de libération hypothalamiques vont atteindre les cellules glandulaires du lobe antérieur de l’hypophyse. Le noyau arqué, situé à la base de la région périventriculaire de l’hypothalamus, est un exemple de région hypothalamique possédant plusieurs sous-populations de ces neurones parvocellulaires ayant une fonction neuroendocrine. Les axones de ces neurones, situés surtout dans la région ventrolatérale du noyau arqué, forment ce qu’on appelle la voie tubéro-infundibulaire qui déverse de la dopamine dans le système porte hypothalamo-hypophysaire. Des cellules glandulaires hypophysaires qui possèdent des récepteurs à la dopamine vont alors diminuer leur libération de prolactine, une hormone qui a plusieurs effets : croissance des glandes mammaires et production du lait chez la femme, sensation de bien-être après l’orgasme chez les deux sexes. Comme les neurones dopaminergiques du noyau arqué sont eux-mêmes inhibés par la succion du bébé qui tète, l’inhibition d’un effet inhibiteur sur les cellules hypophysaires de prolactine va produire une augmentation de la production de celle-ci, et donc plus de lait pour le bébé (voir l’encadré sur les boucles de rétroaction).
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
|
|
|
|
|