Le noyau du faisceau solitaire,
dans le bulbe rachidien, intervient aussi dans l’endormissement par ses
projections sur l’aire
préoptique de l’hypothalamus. On sait par exemple que la stimulation
des nerfs pneumogastriques endort. De plus, ceux qui pratiquent des sports de
combat connaissent bien les effets sur la vigilance des coups portés à
la hauteur du cou. Même chose pour les masseurs de Bali qui connaissent
les effets relaxant des massages de la région carotidienne. Le terme carotide
lui-même vient d’ailleurs du mot grec signifiant « qui
provoque un sommeil profond ».
À la lumière de ces
circuits complexes du sommeil, on voit queles insomnies peuvent avoir
plusieurs origines : une persistance des stimulations éveillantes;
un hypofonctionnement du système anti-éveil; ou encore un retard
de phase de l'horloge.
LES INTERRUPTEURS NEURONAUX DE L'ÉVEIL
ET DU SOMMEIL
Une fois activé,
le réseau
de l'éveil est entretenu par des stimulations internes et externes.
Comment
nous vient alors l'envie de dormir ? Ces stimulations doivent d’abord bien
sûr diminuer. Mais ensuite, I'endormissement est tout simplement le résultat
d'un mécanisme généré par l'éveil lui-même
!
Ces neurones GABAergiques, qui reçoivent
souvent l’appellation de « sommeil lent – On »,
ont leur maximum d’activité durant le sommeil lent et sont inactifs
durant l’éveil et le sommeil paradoxal. La stimulation électrique
de ces neurones induit rapidement le sommeil et leur destruction provoquent des
insomnies. Insomnie toutefois interrompue par l’injection d’un analogue
du GABA (le muscimol) dans l’hypothalamus postérieur, où convergent
plusieurs éléments du système d’éveil.
La localisation d’un tel système favorisant
l’endormissement dans l’aire préoptique de l’hypothalamus
est idéale puisqu’il s’agit d’un carrefour stratégique
qui contrôle des fonctions vitales comme la thermorégulation, la
faim, la reproduction, etc. L’aire préoptique serait donc en mesure
d’analyser l’état fonctionnel de l’organisme afin de
déclencher le sommeil avant que la fatigue ne soit trop intense, au moment
idéal indiqué par l’horloge biologique.
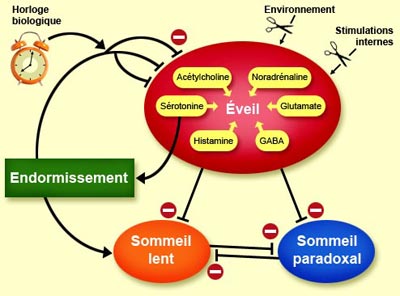
Le
noyau suprachiasmatique, principale structure de cette horloge biologique,
participe donc aussi au déclenchement du sommeil. Lorsque ses neurones
sont lésés, les longues phases d’éveil raccourcissent,
et leur répartition au cours d’une journée devient aléatoire.
Les neurones du noyau suprachiasmatique influencent l’éveil par l’entremise
d’un de leurs neuropeptides : la vasopressine. À
noter que les effets cérébraux de la vasopressine synthétisée
par le noyau supraschiasmatique sont totalement différents de ceux de la
vasopressine produite par l’hypophyse postérieure, qui agit notamment
sur le rein et la pression artérielle.
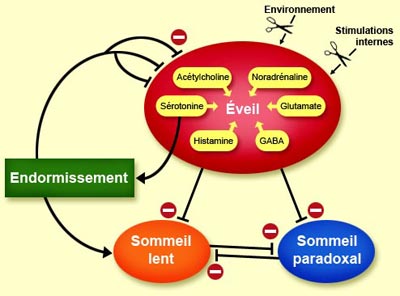
Mais si l’on revient à la sérotonine,
celle-ci joue donc un double jeu spécifique : d’une part, elle
est produite massivement durant l’éveil et contribue de façon
importante à cet état; mais d’autre part, elle a un rôle
fondamental pour l’endormissement qui mène au sommeil lent. Ce paradoxe
a été long à élucider. La sérotonine
a même été longtemps considérée comme
« l’hormone du sommeil », car la lésion des
neurones qui la produisent ou l’inhibition de sa synthèse provoque
une insomnie de plusieurs jours. On pouvait aussi restaurer le sommeil chez un
animal rendu insomniaque par absence de sérotonine en lui injectant le
précurseur immédiat de la sérotonine dans l’aire préoptique
de l’hypothalamus antérieur.
On comprend
aujourd’hui beaucoup mieux pourquoi le manque de sérotonine dans
l’hypothalamus antérieur empêche la venue du sommeil. Cela
a d’ailleurs mené à une conception de l’aire préoptique
de l’hypothalamus antérieur non pas comme un centre du sommeil, mais
plutôt comme une région exerçant une inhibition de l'éveil.
Conception qui s’est vue par la suite confirmée au niveau électrophysiologique.
L'activité unitaire des neurones du raphé est en effet maximale
pendant l'éveil, diminue au début du sommeil et s'arrête pendant
le sommeil paradoxal. Ce silence électrique qui s’installe progressivement
à partir du sommeil lent jusqu’au sommeil paradoxal est associé
à l'arrêt de la libération synaptique de la sérotonine.
L’éveil ayant été contré, la sérotonine
a fait son travail et peut s’estomper.
Les
cellules pacemakers du sommeil lent peuvent donc s’exprimer grâce
à l'inhibition du réseau de l'éveil par le système
anti-éveil. Parallèlement, la "desinhibition"
du pacemaker thalamique contribue aussi à son avènement. L'activité
rythmique qui se met alors en marche dans le thalamus empêche le cortex
d'effectuer les processus cognitifs qui nécessitent une activité
communication thalamo-corticale rapide comme celle qui se déroule lors
de l'éveil ou du rêve.
Chacun
de nos différents états de vigilance (veille, sommeil lent, sommeil
paradoxal) a un
rythme d’oscillation qui lui est propre. Cette rythmicité est
le fruit des interactions entre le thalamus et le cortex, qui elles-mêmes
dépendent des modulations du tronc cérébral et de l’hypothalamus.
Par
exemple, les neurones de la voie ascendante, situés dans la partie rostrale
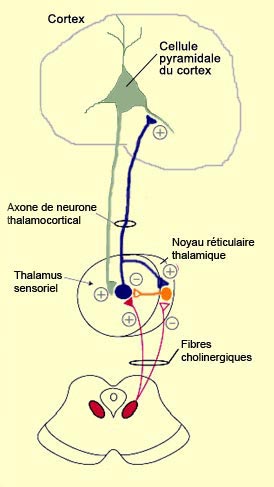
de la protubérance, envoient leurs axones jusqu’au thalamus avec
lequel ils font des connexions cholinergiques. Ces neurones des noyaux
mésopontins cholinergiques innervent les aires sensorielles du thalamus
mais aussi son noyau réticulaire, une couche de neurones
qui enrobe le thalamus comme une pelure. Ce noyau réticulaire thalamique
(qui malgré son nom n’a rien à voir avec la formation réticulée)
a une influence inhibitrice globale sur le thalamus par l’entremise du neurotransmetteur
GABA.
Comment se fait-il alors que ces deux noyaux aux effets opposés
reçoivent la même innervation cholinergique ? C’est que leur
réponse va être différente parce qu’ils n’ont
pas le même type de récepteur : le thalamus sensoriel sera sensibilisé
par l’activation de récepteurs nicotiniques à
l’acétylcholine; tandis que le thalamus réticulaire sera inhibé
par l’activation de récepteurs muscariniques.
Quand le cerveau est éveillé, ses réseaux cholinergiques,
histaminergique et noradrénergiquesactivent donc doublement
le thalamus : une activation directe par facilitation du thalamus sensoriel;
et une activation indirecte, par inhibition de noyau réticulaire inhibiteur.
Il est important de remarquer que l’acétylcholine
n’excite pas directement les neurones thalamiques sensoriels, se contentant
plutôt de les sensibiliser en les dépolarisant légèrement.
Cette dépolarisation fait passer leur pattern de décharge en bouffée
à une décharge régulière de potentiels
d’action. En étant ainsi dépolarisés, les neurones
thalamiques deviennent plus sensibles aux afférences sensorielles. Les
neurones corticaux, qui reçoivent d’importantes connexions de ces
neurones thalamocorticaux vont ainsi voir leur activité désynchronisée
et le tracé de l’EEG devenir typique de l’éveil, c’est-à-dire
de faible amplitude mais de haute fréquence. À noter que les cellules
pyramidales du cortex reçoivent également une excitation cholinergique
nicotinique directe du
noyau basal de Meynert en plus de celle des neurones thalamocorticaux.
Puis,
durant les minutes où se déroule l’endormissement, on observe
une baisse dans la fréquence de décharge des neurones à noradrénaline,
à acétylcholine et à sérotonine du système
activateur du tronc cérébral. Le thalamus est par conséquent
moins activé.
Parallèlement, l’inhibition
sur le noyau réticulaire du thalamus diminue tout autant et son effet inhibiteur
sur les neurones thalamo-corticaux va se faire de plus en plus sentir. Car cette
levée des influences inhibitrices permet aux neurones réticulaires
d’exprimer à nouveau leur activité spontanée oscillatoire.
Les potentiels d’action périodiques qu’émettent alors
les neurones « pacemaker » GABAergiques du noyau réticulaire
vont
entraîner une hyperpolarisation cyclique des neurones thalamo-corticaux,
contribuant ainsi à la génération de l’activité
rythmique du thalamus. Celui-ci devient alors de plus en plus insensible aux stimuli
de l’environnement, ce qui est le propre des stades profonds du sommeil
lent.
Avec la phase II du sommeil lent, l’activité corticale
devient sous l'influence d'une activité automatique d'origine thalamique
caractérisée par les
fuseaux du sommeil sur l’EEG. Ceux-ci proviennent, comme on vient de
le décrire, des décharges rythmiques des neurones réticulaires
qui produisent chez les neurones thalamocorticaux des hyperpolarisations cycliques
suivies de bouffées de potentiels qui, transmises aux cellules corticales,
y génèrent les fuseaux.
Quant aux ondes lentes de grande
amplitude produites durant les stades III et IV du sommeil lent, elles résultent
de l’hyperpolarisation des cellules pyramidales du néocortex déclenchée
par des interneurones GABAergiques locaux, vraisemblablement sous l’influence
des neurones préoptiques de
l'hypothalamus antérieur. Les neurones thalamiques, dont le potentiel
de membrane est alors encore plus négatif que pendant les fuseaux du
sommeil (observables surtout au stade II), contribueraient aussi à ces
ondes lentes corticales.
Enfin, lors du sommeil paradoxal,
l’EEG désynchronisé qui caractérise cet état
est produit entre autre par une influence cholinergique sur les cellules thalamiques
qui empêche l’expression de son activité oscillatoire rythmique
par les mêmes mécanismes décrits plus haut pour l’éveil.
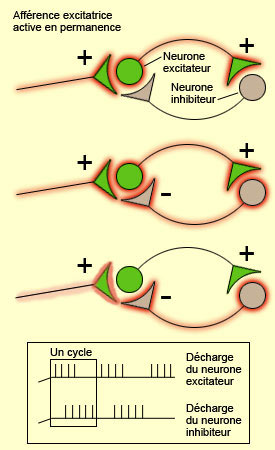
Différents
types de connectivité neuronale favorise l’émergence de bouffées
rythmiques de potentiels d'action au sein d’un réseau de neurones.
Ainsi, l’un des plus simple que l’on peut identifier est la connexion
réciproque entre un neurone excitateur et un neurone inhibiteur, lesquels
sont activés par un troisième neurone (dont le pattern d’activation
peut être, lui, régulier et sans bouffées rythmiques). Aussi
longtemps que dure cette activation constante du neurone excitateur en provenance
de l’extérieur, l’activité de ce neurone excitateur
va être périodiquement interrompue parce que celui-ci active le neurone
inhibiteur qui l’inhibe en retour. L’arrêt temporaire de l’activité
du neurone excitateur fait immédiatement cesser celle du neurone inhibiteur,
ce qui rend à nouveau le neurone excitateur réceptif à l’activation
constante de l’afférence extérieure. D’où l’activation
du neurone excitateur qui reprend, entraînant rapidement celle du neurone
inhibiteur, et la reprise du cycle oscillatoire.