Les
dendrites sont également émis par le neurone afin
de recevoir éventuellement des contacts en provenance d'autres neurones.
Plus courts que l'axone et très ramifiés, ces prolongements grandissent
néanmoins en développant eux aussi un cône de croissance à
leur extrémité.
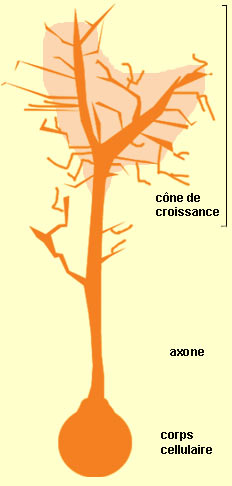
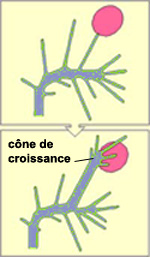
LE CÔNE DE CROISSANCE
Un des aspects les plus extraordinaires
du développement du système nerveux est sans doute la capacité
des axones en croissance à trouver leurs cellules cibles. Celles-ci sont
situées souvent à des millimètres ou même à
des centimètres plus loin, ce qui constitue de très grandes distances
à cette échelle. Cette capacité leur vient du cône
de croissance, une structure située à l’extrémité
de l’axone qui s’allonge.
Le cône de croissance est
formé de divers
prolongements qui avancent et reculent à la recherche de signaux de
guidage, un peu comme des doigts qui s’étendent et se rétractent
pour palper ce qu’il y a aux alentours. C’est donc en interagissant
avec son environnement que le cône de croissance trouve des signaux qui
le guideront à l’endroit où il doit établir
des connexions avec d'autres neurones.
Ces signaux de guidage sont
des molécules qui renseignent le cône
de croissance sur la direction à suivre. Elles sont parfois fixées
directement au substrat sur lequel se déplace le cône de croissance.
Les molécules signaux peuvent aussi être sécrétées
par une cellule et diffuser librement dans le milieu environnant. Il se forme
ainsi un gradient qui influence à distance la trajectoire de l’axone
en développement.
Si le cône de croissance peut être
influencé par ces molécules, c’est qu’il possède
des récepteurs spéciaux capables de les détecter. C’est
donc grâce au déploiement de molécules guides et à
leurs récepteurs spécifiques répartis sur différents
neurones que les grandes voies neuronales se mettent en place dans l’embryon.
Il y a plus de 100 milliards de
neurones dans le cerveau humain et chacun fait plusieurs milliers de synapses
avec d'autres neurones. Les combinaisons possibles excèdent donc de beaucoup
les quelques 20 ou 30 000 gènes que nous possédons. Ce caractère
limité de l'information génétique suggère que d'autres
facteurs extrinsèques comme les interactions entre cellules ou les molécules
chimioattractives jouent un rôle important dans le développement
du système nerveux.
LES MOLÉCULES QUI GUIDENT LE CÔNE DE CROISSANCE
Le cône
de croissance qui guide l’axone vers la cellule avec laquelle il doit
former une synapse est un peu comme un automobiliste
qui circulerait dans un pays inconnu sans carte routière. Il doit s’en
remettre uniquement aux panneaux de signalisation qu’il rencontre sur sa
route. Pour le cône de croissance, ces signaux routiers prennent la forme
de molécules. Celles-ci peuvent être divisées en deux grandes
familles.
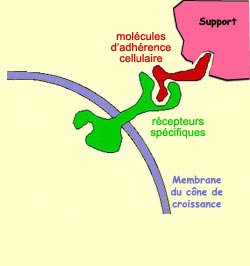
La
première est faite de molécules attachées à différents
supports situés sur la route qu’emprunte le cône de croissance.
Comme des panneaux de signalisation routière que l’on reconnaît
sur le bord de la route, ces molécules d’adhérence cellulaire
sont reconnues par des récepteurs
spécifiques situés sur la membrane du cône de
croissance.
C’est donc par contact direct avec ces molécules
que d’autres signaux sont transmis à l’intérieur de
l’axone en croissance, des signaux qui en bout de ligne orientent la direction
de sa croissance. Par opposition à la famille suivante, on qualifie ces
molécules de non diffusibles.
Source:
Dr. Brian E. Staveley Department of Biology Memorial University of Newfoundland
Changement de direction d’un cône de croissance ayant touché
un substrat avec des molécules d’adhésion compatibles.
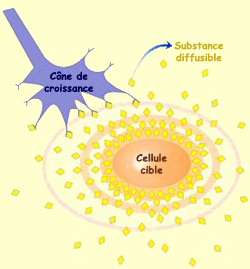
La seconde famille fait donc intervenir des molécules
non pas fixées sur un substrat, mais libres de diffuser dans le milieu
aqueux qui entoure le cône de croissance. Ce mécanisme reçoit
l’appellation de chimiotropisme.
Transposé à l’échelle de l’individu, le chimiotropisme
pourrait correspondre à l’odeur de café fraîchement
torréfié qui attire irrémédiablement l’amateur
d’espresso vers le lieu de la torréfaction. Dans ce cas, on parlera
de chimioattraction. Mais il y a aussi des molécules qui ont un effet de
chimiorépulsion sur l’axone en croissance. La mauvaise odeur d’un
dépotoir en serait l’équivalent sur la trajectoire d’un
marcheur…
Finalement il existe une troisième catégorie de molécules
qui, sans être des signaux de signalisation en tant que tel, sont néanmoins
nécessaires à l’allongement de l’axone. On les appelle
les facteurs de croissance
et ils jouent un rôle crucial dans la
formation des connexions synaptiques.
En plus de permettre la
survie de certains neurones particuliers durant le développement
et de participer à l’organisation de leurs connexions initiales,
la compétition pour les molécules trophiques permet aux ramifications
des neurones et à leur connexions de se modifier tout au long de la vie
suite aux changements de l’activité nerveuse provoqués par
les processus
d’apprentissage.
FACTEURS DE CROISSANCE ET MORT NEURONALE
À partir du moment
où les neurones commencent à se constituer en circuits, il se produit
un changement d’échelle dans le développement du système
nerveux : de cellules isolées, nous passons à un réseau de
milliers d’éléments interconnectés. C’est ce
réseau qui acquiert la capacité de traitement de l’information
qui fait du cerveau l’outil puissant que l’on connaît.
Mais ce réseau est loin d’être parfait au départ.
On assiste en particulier à une production de deux ou trois fois plus de
neurones que nécessaire, suivie par une réduction subséquente
des neurones excédentaires. Quels sont donc les neurones qui vont survivre
et pourquoi ceux-là ?
On sait maintenant que la survie d’un
neurone dépend en grande partie de la relation qu’il va entretenir
avec sa cellule cible. Des expériences ont par exemple montré qu’en
réduisant le nombre de cellules cibles, on réduit le nombre de neurones
qui doivent venir les connecter. À l’opposé, l’existence
d’une plus grande population cellulaire à innerver maintient en vie
un plus grand nombre de motoneurones.
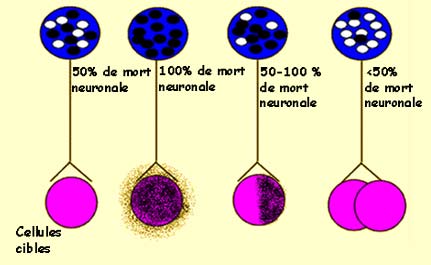
Le taux de survie des neurones dépend de l’importance
de la population des cellules cibles qu’ils innervent. Les zones ombragées
symbolisent la destruction de cellules cibles.
Le
mécanisme en jeu pour expliquer ce phénomène implique donc
une certaine compétition entre les neurones pour des facteurs de survie
particuliers appelés facteurs de croissance ou facteurs
trophique. Du grec trophè, pour nourriture, ces substances ne
sont cependant pas des sources d’énergie comme le
glucose ou l’ATP, mais bien des
molécules sécrétées par les cellules cibles qui
sont indispensables à la survie et à la croissance des neurones.
Pour reprendre la métaphore automobile,
il ne s’agit pas ici de l’essence qui sert de source d’énergie
au véhicule mais à quelque chose de l’ordre du permis de conduire.
En effet, son absence amène de gros problèmes au développement
de l’axone et bien souvent son élimination pur et simple des routes
neuronales…
La mort de ces neurones sera alors bien différente
de celle provoquée par une lésion ou une maladie. Il s’agira
plutôt d’une disparition en douceur et programmée, appelée
apoptose.
L’apoptose implique en effet l’expression
d’une multitude de gènes spécifiques qui entraînent
la dégénérescence cellulaire d’une façon qui
n’est pas dommageable pour l’ensemble de l’organisme. Ces gènes
sont d’ailleurs souvent aussi impliqués dans la différenciation
et le contrôle du cycle cellulaire normal (voir capsule outil à gauche).
Dans le cerveau d’un bébé,
les neurones reçoivent autour d’une fois et demie plus de synapses
que les neurones de cerveaux adultes. Ces nombreuses connexions se maintiendraient
relativement constantes jusqu’à la puberté, mais à
l’adolescence, une perte marquée se produirait : les neurones du
cortex visuel primaire perdraient alors en moyenne 5 000 synapses par seconde
!
Quand une cellule cible perd des
connexions d’autres neurones pour ne conserver celles en provenance que
d’un seul neurone, on parle souvent à tort «d’élimination
de synapses». Pour être juste, on devrait dire qu’il s’agit
plutôt d’une réduction du nombre d’afférences
différentes que reçoit la cellule cible, le nombre total de synapses
ne faisant généralement qu’augmenter au cours du développement.
FORMATION ET STABILISATION SÉLECTIVE DES SYNAPSES
Il arrive souvent que le
guidage des axones jusqu’au voisinage de leur cible soit le même
pour des neurones impliqués dans différents circuits. Comment chaque
axone reconnaît-il alors la bonne cellule cible ? Dans certains cas, il
semble que des molécules semblables aux molécules
d’adhérence cellulaire agissent comme des étiquettes qui
permettent aux différents cônes
de croissance des axones de reconnaître les bonnes cellules cibles.
De plus, l’endroit exact où se forme la
synapse sur la cellule est étroitement contrôlé par un
ensemble de molécules. Car la synapse exige la présence de structures
moléculaires particulières pour fonctionner convenablement.
Ainsi,
au niveau de la
jonction neuro-musculaire où la formation des synapses a été
la plus étudiée, on appelle « zone active » cet endroit
où la machinerie de libération des neurotransmetteurs des axones
s’aligne avec des regroupements de forte densité de récepteurs
à l’acétycholine dans la fibre musculaire.
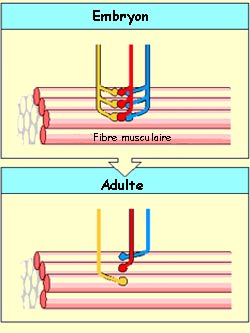
Une
fibre musculaire peut, dans un premier temps, recevoir des connexions en provenance
de plusieurs motoneurones. Mais elle les perdra progressivement pour ne conserver
qu’une connexion d’un seul motoneurone.
On
a pu montrer que c’est l’activité électrique de la fibre
musculaire qui régule ce processus. Plus la fibre est active, plus les
synapses s’éliminent rapidement, à l’exception de celles
provenant d’un seul motoneurone. À l’opposé, réduire
l’activité musculaire retarde cette sélection.
Il
existe de nombreuses preuves que de telles réorganisations synaptiques
interviennent également dans le cerveau immature. Un neurone peut autant
perdre des connexions initialement établies avec d’autres neurones,
ou encore voir ses connexions avec certains neurones se multiplier. Et encore
une fois, c’est l’activité nerveuse qui permet le maintient
ou l’augmentation des contacts synaptiques alors que l’absence de
stimulation amène l’élimination de ces synapses inutiles.
C’est pourquoi l’on dit que la
stabilisation de nos synapses est sélective et que c’est l’activité
des circuits qui est à l’origine de cette sélection.
Le câblage du cerveau d’un
individu résulte d’un plan génétique initial qui laisse
par la suite les connexions neuronales se remodeler au fil des interactions avec
l’environnement. Deux grands patterns de circulation de l’information
se dessinent alors : la convergence, c’est-à-dire
plusieurs fibres nerveuses qui arrivent sur la même cellule cible, et la
divergence, soit un même neurone qui contacte différentes
cellules.