Un livre accompagne le site depuis octobre 2024. Découvrez-le ici >
De l'embryon à la morale
Dans le système nerveux central,
les cônes de croissance des nerfs optiques changent de forme lorsqu’ils
arrivent au chiasma
optique. Les cônes de croissance des axones des cellules
ganglionnaires de la rétine ralentissent leur avancée
et prennent une forme plus complexe au moment de « choisir » de croiser
ou non le chiasma.
Le même phénomène
est observé en périphérie avec les cônes de croissance
des motoneurones qui se mettent à chercher davantage quand ils entrent
dans les ébauches musculaires des membres en développement.
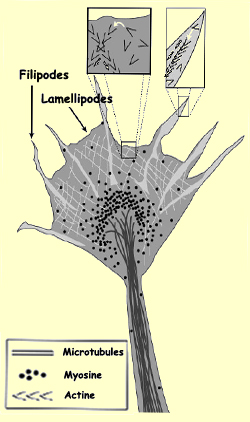
LE CÔNE DE CROISSANCE
Quand
les neuroblastes
ont terminé leur
migration ou même pendant celle-ci, ils émettent des prolongements
appelés neurites
qui s’allongent par leur extrémité. L’un de ces neurites,
qui deviendra l’axone, devra d’ailleurs croître sur de longues
distances avant d’atteindre sa cible. Son allongement sera rendu possible
grâce à une structure située à son extrémité,
le cône de croissance.
Le cône de croissance
de l’axone ou des dendrites est fait de feuillets membranaires aplatis,
les lamellipodes, desquels partent de fines expansions appelées
filipodes qui s’étirent et se rétractent
constamment pour explorer l’environnement. Quand un filipode, au lieu de
se rétracter, s’accroche au substrat, il fait avancer le cône
de croissance dans cette direction.
Les cônes de croissance d’axones
qui progressent le long d’une voie déjà frayée par
d’autres axones ont une forme plutôt simplifiée. Par contre,
quand l’axone s’aventure dans une nouvelle voie ou lorsqu’il
arrive à une intersection où il doit choisir sa direction, son
cône de croissance se complexifie de façon spectaculaire : il s’aplatit
et émet de nombreux filipodes pour chercher activement les indices susceptibles
de le guider (voir encadré).
L’actine
est représentée en gris pâle sur le dessin principal et par
des >>>>> dans les agrandissements. Les flèches blanches
des agrandissements représentent la polimérisation de l’actine.
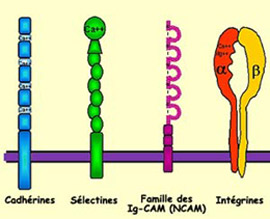
L’une des caractéristiques
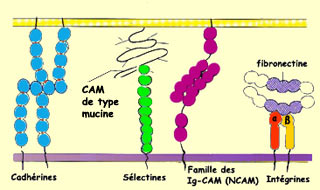
de pratiquement toutes les CAM est de posséder, dans leur partie extracellulaire,
des motifs répétitifs d’acides aminés. On distingue
cependant deux familles de CAM selon qu’elles nécessitent ou pas
la présence de calcium pour adhérer : les cadhérines
en ont besoin alors que les NCAM (pour « neural cell adhesion
molecules », en anglais) sont indépendantes du calcium.
LES MOLÉCULES QUI GUIDENT LE CÔNE DE CROISSANCE
Parmi les
différents signaux qui guident le cône de croissance, certains
sont dits non diffusibles. Il s’agit de molécules intervenant dans
les contacts de cellule à cellule, comme le vaste groupe des molécules
d’adhérence cellulaire (ou CAM pour «
cell adhesion molecules », en anglais).
Les
CAM sont des protéines transmembranaires qui font sailli à la surface
des cellules. Elles peuvent ainsi interagir avec des récepteurs
spécifiques aux CAM situés sur les cônes de croissance.
La reconnaissance entre une CAM et son récepteur déclenche
alors une cascade biochimique de seconds messagers dans le cône de croissance,
cascade qui va aboutir à l’activation d’enzymes (kinases, phosphatases,
protéases, etc.) dont les effets vont contribuer à l’élongation
de l’axone.
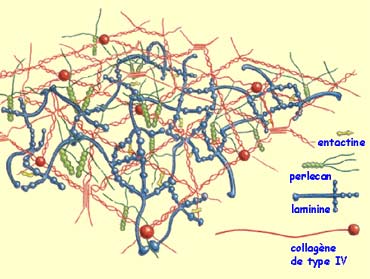
Des récepteurs
des cônes de croissance sont aussi sensibles à des protéines
situées non pas sur la membrane des cellules mais distribuées dans
la matrice extracellulaire, agglomérat de substances produites par la cellule
mais non directement fixées à elles.
Les plus connues de
ces molécules d’adhérence de la matrice extracellulaire sont
les laminines, les collagènes et la fibronectine.
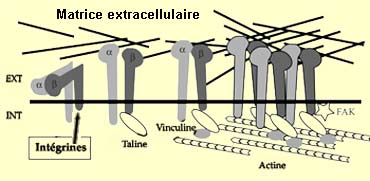
Une catégorie de récepteurs du cône de croissance appelée
intégrines reconnaît spécifiquement ces molécules.
Outre
les protéines de surface, une autre catégorie importante de molécule
peut influencer la direction de l’allongement de l’axone. Il s’agit
de substances sécrétées par les cellules cibles et qui vont
se diffuser dans le milieu extracellulaire environnant. Ces substances, émises
en très petites quantités peuvent être soit
chimioattractives et attirer l’axone, soit chimiorépulsives et le
repousser.
Ces facteurs dits chimiotropes diffèrent
cependant d’une autre catégorie de molécules diffusibles appelées
facteurs
trophiques. Ces derniers, dont fait partie le NGF (« Nerve
Growth Factor »), ont pour fonction de maintenir le neurone en vie et faciliter
la croissance de ses prolongements.
Le BDNF est particulièrement
important pour la survie des neurones
corticaux des aires visuelles notamment. La plupart des récepteurs
sur lesquels se fixent les neurotrophines, appelés récepteur trk,
sont des protéines kinases qui phosphorylent des résidus tyrosine
situés sur d’autres protéines qui sont leur substrat. En d’autres
termes, elles ajoutent des atomes de phosphore à l’acide aminé
tyrosine de certaines protéines pour en modifier la forme et par le fait
même la fonction. Dans le cas qui nous intéresse ici, cette phosphorylation
va éventuellement avoir un effet sur l’expression génique.
Il semble que les neurotrophines,
et en particulier le BDNF qui est très largement exprimé dans le
SNC, jouent un rôle non négligeable dans la
plasticité synaptique, c’est-à-dire dans
les modifications morphologiques et physiologiques des synapses en réponse
à des changements d’activité neuronale. On sait par exemple
que la synthèse et la libération du BDNF par les neurones centraux
sont sous le contrôle de l’activité neuronale ce qui permet
à ces neurotrophines de moduler les transmissions GABAergiques et glutamatergiques
de certaines structures cérébrales comme l’hippocampe
et le cortex
visuel.
FACTEURS DE
CROISSANCE ET MORT NEURONALE
Bien qu’ils contribuent
au développement et au maintien de nos réseaux de neurones, les
facteurs trophiques (ou facteurs de croissance)
diffèrent des molécules
de guidage de l’axone. Il s’agit d’une autre catégorie
de molécules qui sont sécrétées par les cellules cibles
dont le rôle n’est pas d’aider l’axone à s’orienter
mais d’assurer sa survie une fois qu’il a formé certaines connexions
synaptiques fonctionnelles.
Les facteurs trophiques sont émis
en quantité limitée par les cellules cibles, de sorte que seul un
sous-ensemble des neurones les innervant va en recevoir assez pour survivre. Autrement
dit, les neurones ont besoin d’une quantité minimale de facteur trophique
pour survivre et tout porte à croire qu’il y a compétition
entre les neurones pour les facteurs trophiques disponibles. Ceux qui en manquent
disparaissent tout simplement par apoptose, le processus de mort cellulaire programmé
de l’organisme (voir la capsule outil à gauche).
Le premier
facteur trophique à avoir été découvert est le facteur
de croissance des nerfs ou NGF (pour « Nerve Growth
Factor », en anglais). Le NGF a été identifié comme
étant une protéine constituée de trois sous-unités,
dont l'une surtout est réellement indispensable à la protection
des neurones (la sous-unité ß, elle-même formée de deux
molécules identiques de 118 acides
aminés).
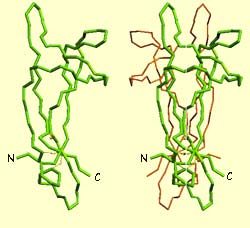
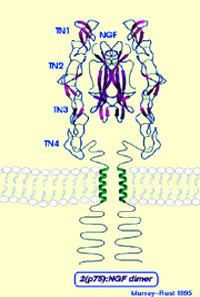
L’image de gauche montre la
structure moléculaire du NGF qui est formée de deux parties homologues
accolées l’une contre l’autre selon leur axe le plus long.
L’image de droite montre la molécule de NGF fixée au cœur
d’un de ses récepteurs.
Bien
que le NGF soit le prototype et sans doute le plus étudié des facteurs
trophiques, il n’affecte que certaines catégories de neurones périphériques.
Depuis le milieu des années1980, plusieurs recherches ont mis en évidence
d’autres facteurs trophiques apparentés. On appelle maintenant cette
famille de molécules les neurotrophines. Elles comptent,
outre le NGF, trois autres molécules bien caractérisées :
le facteur neurotrophique dérivé du cerveau ou BDNF
(pour « Brain-Derived Neurotrophic Factor », en anglais), la neurotrophine-3
(NT-3) et la neurotrophine-4/5 (NT-4/5).
Des expériences ont en effet démontré que la
croissance des neurites pouvait être contrôlée localement
par les facteurs de croissance, sans nécessairement passer par la machinerie
enzymatique située dans le corps cellulaire. Par conséquent, un
neurone peut voir certaines de ses ramifications s’étendre alors
qu’en même temps d’autres se contractent, ce qui s’observe
effectivement lors de la formation des synapses.
L’affinité entre un
axone et sa cible est un peu comme les codes de couleurs dans les câbles
électriques formés de plusieurs fils qui permettent de raccorder
les bons fils ensemble.
Mais contrairement à l’exemple
de ces codes de couleurs mutuellement exclusifs, des travaux ont montré
que l’affinité entre un neurone et sa cible n’est pas très
sélective. On observe plutôt une préférence de certains
axones pour certaines cellules cibles qui n’empêche pas l’axone
d’établir aussi des contacts synaptiques avec d’autres neurones.
Les associations entre les neurones et leur cible se font donc plutôt selon
un continuum de préférence. Ce continuum va de l’impossibilité
absolue de faire des connexions avec des cellules gliales par exemple, à
la possibilité de contacter indifféremment telle ou telle cellule
d’une population donnée.
Durant la formation des synapses
de la plaque motrice, même les propriétés biologiques du récepteur
nicotinique de l’acétylcholine changent. Son taux de remplacement
diminue, sa capacité à laisser passer les ions augmente et la combinaison
des sous-unités qui le compose est modifiée. Tous ces changements
contribuent à l’ajustement fin de la jonction
neuromusculaire.
FORMATION ET STABILISATION SÉLECTIVE DES SYNAPSES
La synapse n’est pas
immédiatement fonctionnelle quand le cône de croissance fait contact
avec sa cible. La formation d’une synapse est un processus graduel qui a
été beaucoup étudié à la jonction neuromusculaire.
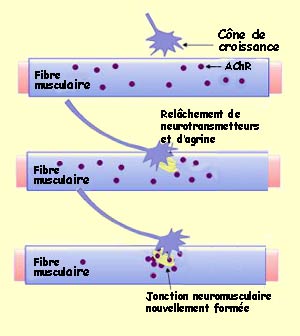
On sait par exemple que le cône de croissance du motoneurone
émet de l’acétylcholine spontanément, avant même
d’atteindre la fibre musculaire. D’autre part, des récepteurs
nicotiniques de l’acétylcholine sont répartis uniformément
sur la membrane de la fibre musculaire. Mais peu de temps après la formation
du contact entre l’axone et la fibre musculaire, les récepteurs nicotiniques
s’accumulent rapidement à l’endroit de la future synapse alors
que la population de récepteurs extra-synaptiques diminue de manière
importante.
L’un
des indices moléculaires qui guide la formation de ces connexions si précisément
ajustées l’une à l’autre est une molécule nommée
agrine. L’agrine est synthétisée par le corps
cellulaire du neurone présynaptique, transportée le long de l’axone
et libérée par les fibres nerveuses en croissance. Elle se lie alors
à des
récepteurs post-synaptiques dont l’activation permet le regroupement
des récepteurs de l’acétylcholine.
En plus de
cette redistribution des récepteurs, de nouveaux récepteurs sont
insérés dans la membrane vis-à-vis la synapse. Le corollaire
de cette observation est qu’il y a synthèse d’ARN
messagers de ces récepteurs dans le noyau le plus près de la
jonction neuromusculaire.
Le
développement des voies neuronales se fait initialement sous la gouverne
de mécanismes génétiquement programmés. Mais ces mécanismes
intrinsèques ne peuvent former que des circuits esquissés grossièrement
faits d’une multitude de synapses extrêmement redondantes. Un processus
de sélection est donc nécessaire pour diminuer le nombre ces synapses
et raffiner les circuits nerveux.
Ce processus de sélection est
dépendant de l’activité des neurones : c’est l’expérience
sensori-motrice de l’individu qui va permettre de valider le câblage
initial et d’ajuster l’organisation fine des réseaux de neurones.
La question qui se pose est donc de savoir comment l’activité des
neurones réagissant à l’environnement peut-elle affecter le
développement des circuits nerveux ?
Pour y répondre,
on doit retourner au
postulat de Hebb, formulé à l’origine pour expliquer les
bases cellulaires de l’apprentissage
et de la mémoire. Celui-ci s’avère également applicable
aux modifications synaptiques importantes qui surviennent durant le développement
du système nerveux. Hebb propose que l’activité corrélée
entre deux neurones fait en sorte qu’une synapse se renforce. Appliquée
au développement, on dira que si deux neurones reliés à la
même cible émettent des signaux coordonnés, leurs connexions
sont renforcées. Au contraire, si ces signaux sont déphasés,
les synapses sont affaiblies.
Par conséquent, les terminaisons
synaptiques dont l’activité durant le développement ne serait
que rarement corrélée à celle du neurone postsynaptique finiraient
par s’affaiblir progressivement jusqu’à leur élimination
complète. C’est ce que Changeux et Danchin ont appelé en 1976,
la stabilisation sélective des synapses en montrant que
seules seront conservées les connexions synaptiques intégrées
dans un circuit neuronal fonctionnel.
Alors que la
mort cellulaire permet d’établir une correspondance entre le
nombre de neurones et de leurs cellules cibles, l’élimination de
synapses sert donc plutôt à rendre le pattern d’innervation
plus précis.
Durant toute notre vie, mais plus
particulièrement durant l’enfance, nos connexions synaptiques sont
sculptées par notre expérience sensorielle. Les neurones peuvent,
d’une part, améliorer l’efficacité de leurs connexions
grâce au mécanisme de la PLT
ou, d’autre part, diminuer leur efficacité synaptique par le processus
de DLT.
Bien que ces deux mécanismes contribuent à l’ajustement fin
de nos connexions neuronales, la DLT semble jouer un rôle particulièrement
important dans l’élimination sélective des synapses qui caractérise
certaines périodes
critiques de notre développement.
La DLT induit une
diminution de récepteurs post-synaptiques, ce qui diminuerait l’activité
de ces synapses et pourrait mener leur élimination progressive observée
durant les périodes critiques.