Même au repos, la plupart
de nos muscles sont dans un état de contraction partielle appelé
« tonus musculaire ». Le tonus est maintenu par l’activation
constante d’une petite quantité d’unités motrices qui
se contracte en alternance.
Les fibres musculaires appartenant
à une même unité
motrice ne sont pas situées côte à côte mais plutôt
dispersées dans le muscle. Cette organisation s’explique sur le plan
fonctionnel si l’on prend en considération l’apport énergétique
nécessaire à la contraction musculaire. Celle-ci tire en effet son
énergie de la combustion du glucose amené aux fibres musculaires
par les capillaires sanguins circulant entre les fibres musculaires. Or comme
toutes les fibres innervées par un motoneurone se contractent en même
temps, leur élargissement comprimerait les capillaires les alimentant en
énergie si elles étaient regroupées au même endroit.
Cette diminution rapide de l’apport énergétique amènerait
donc une fatigue rapide de l’unité motrice, d’où la
distribution éparse de leurs fibres dans le muscle pour éviter ce
problème.
LA JONCTION NEUROMUSCULAIRE ET LA CONTRACTION
DES MUSCLES
La contraction
musculaire d’une unité
motrice est initiée par la libération d’acétylcholine
au niveau de la jonction neuromusculaire.
L’activation
des récepteurs nicotiniques cholinergiques
déclenche un potentiel post-synaptique excitateur au niveau de la plaque
motrice de la fibre musculaire. Si celui-ci atteint un certain seuil, un potentiel
d’action musculaire est généré par les canaux sodiques
dépendants du potentiel de membrane.
Ce potentiel d’action
circule à la surface du sarcolemme puis des tubules T pour provoquer la
libération du calcium stocké dans le réticulum endoplasmique.
Ce calcium diffuse dans les myofibrilles qui sont divisés par les stries
Z en segments appelés sarcomères. Dans
chaque sarcomère, des filaments fins et épais peuvent glisser les
uns sur les autres pour rapprocher les stries Z et réduire ainsi la longueur
du sarcomère lors de la contraction musculaire.
Pour
comprendre comment le calcium va provoquer ce glissement des filaments fins et
épais, il faut considérer la protéine formant principalement
les filaments fins, l’actine, et celle formant majoritairement
les filaments épais, la myosine. Et plus particulièrement
les « têtes » des molécules de myosines qui, en s’associant
à celles de l’actine, provoquent un changement de conformation qui
induit une rotation des têtes de la myosine. C’est cette rotation
qui fera glisser d’un cran les filaments fins le long des filaments épais,
un peu comme un mécanisme de crémaillère. Or en l’absence
de calcium, la myosine ne peut pas interagir avec l’actine car ses sites
de liaison sont occupés par une autre protéine, la troponine.
Le calcium qui arrive suite à un potentiel d’action musculaire
va donc, en se fixant sur la troponine, faire deux choses : 1) exposer les sites
d’interaction de la myosine et 2) modifier la forme d’une autre protéine,
la tropomyosine, qui va pour sa part libérer les sites
de fixation spécifiques de la myosine présents sur la molécule
d'actine. L’actine et la myosine pouvant maintenant interagir, la rotation
des têtes de myosine sur celles d’actine devient possible. La contraction
musculaire va donc avoir lieu tant que le calcium et l’ATP seront disponibles.
L’ATP est entre autre nécessaire à la rupture de la liaison
entre l’actine et la myosine. C’est ce qui explique la rigidité
musculaire d’un cadavre lorsque cet apport d’ATP vient qu’à
manquer.
La quantité de calcium émis par le réticulum
endoplasmique dépend de la fréquence des potentiels d’action
dans la fibre muculaire. À partir de 50 stimuli par seconde, la fréquence
est suffisante pour provoquer une contraction musculaire soutenue appelée
tétanos.
La contraction musculaire prend fin quand cessent les
potentiels d’action et que diminue la concentration de calcium dans les
myofibrilles. Cette diminution est causée par la recapture du calcium dans
le réticulum endoplasmique, un processus actif qui nécessite de
l’ATP. Quand la concentration de calcium revient à la normale, la
fibre musculaire se relaxe et revient à sa position détendue.
La myasthénie est une maladie
liée à un défaut de transmission de l'influx nerveux entre
le nerf et le muscle volontaire. Il s’agit d’une maladie dite auto-immune
parce que certains globules blancs fabriquent, chez ceux qui en souffrent, des
anticorps contre leurs propres récepteurs de l'acétylcholine, ce
qui a pour effet de les détruire. L’efficacité de la jonction
neuromusculaire est donc réduite, ce qui amène une faiblesse musculaire
à l’effort ou, dans les formes graves de la maladie, une force musculaire
réduite en permanence.
Des courants dus à la libération
spontanée de vésicules d'ACh peuvent être enregistrés
au niveau de la jonction neuromusculaire en l'absence de toute stimulation de
l'axone moteur. Ces courants miniatures correspondent à l'ouverture d’environ
1600 récepteurs nicotiniques ouverts par l'ACh. Comme il faut 2 molécules
d'ACh pour ouvrir un seul canal, une vésicule contient alors 3200 molécules
d'ACh. Le pic du courant d’une plaque motrice correspondant à environ
100 courants miniatures, soit à l'ouverture de 160 000 récepteurs
nicotiniques par l'acétylcholine libérée, donc 320 000 molécules
d'ACh.
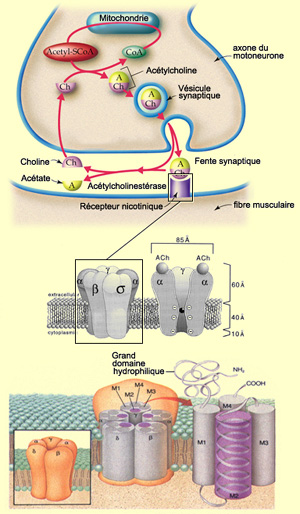
LE RÉCEPTEUR NICOTINIQUE DE L'ACÉTYLCHOLINE
À la jonction
neuromusculaire, l’acétylcholine est synthétisée
dans le cytoplasme de la terminaison nerveuse à partir de l’acétylCoA
et de la choline. Environ la moitié de cette choline vient de la choline
recaptée par la terminaison nerveuse après hydrolyse de l’acétylcholine
par l’acétylcholinestérase dans la fente synaptique. Plusieurs
milliers de molécules d’acétylcholine sont ainsi stockée
par vésicule synaptique (voir encadré).
Dès leur libération,
environ la moitié des molécules d’acétylcholine sont
hydrolysées par l’enzyme acétylcholinestérase. Mais
il s’en accumule bientôt une telle quantité dans la fente synaptique
que l’enzyme ne peut tout dégrader et l’autre moitié
atteint les récepteurs nicotiniques. Tout cela se déroule évidemment
très rapidement puisqu’on estime la demi-vie de l’acétylcholine
à 1 ou 2 millisecondes, l’acétylcholinestérase étant
capable d’hydrolyser 4000 molécules d’ACH par site et par seconde.
La répartition de l’acétylcholinestérase se superpose
d’ailleurs parfaitement à celle des récepteurs nicotiniques
postsynaptiques.
C’est donc principalement une réaction
biochimique qui termine ici la
transmission synaptique, contrairement aux catécholamines où
la diffusion à l’extérieur de la fente synaptique et surtout
la
recapture par le bouton présynaptique en sont les principales responsables.
En ce qui concerne les récepteurs à l’acétylcholine,
on les subdivise en deux grandes familles : nicotiniques
et muscariniques. Ceux de la jonction neuromusculaire, de type nicotiniques,
sont des protéines pentamériques qui forment un canal ionique fiché
dans la couche bi-lipidique de la membrane postsynaptique. On en compte près
de 10 000 au micromètre carré. L’acétylcholine se fixe
sur la partie extra-cellulaire des deux sous-unités alpha de cette protéine
canal qui compte 5
sous-unités en tout.
Chaque sous-unité comprend elle-même
quatre domaines hélicoïdaux transmembranaires identifiés M1
à M4. Dans chaque sous-unité, c’est la partie M2 de ces hélices
qui forment la paroi du canal ionique. Ce canal ionique, ouvert par le changement
de conformation allostérique provoqué par la fixation de l’acétylcholine,
est également perméable à Na et K. La perméabilité
au calcium ne contribue que pour 2,5 % à la perméabilité
totale. La demi-vie des récepteurs nicotiniques adultes est de 4 à
6 jours.