Capsule outil: Le modèle neurodynamique de la perception de Walter J. Freeman
Nos récepteurs sensoriels sont excités de manière variable, et nous percevons pourtant un même objet. Par exemple, en changeant la position de mon œil par rapport à un objet, des cellules réceptrices différentes sont excitées, mais je perçois pourtant l'objet comme invariable. Ce phénomène touche toutes les formes de perception. Ainsi, dans la perception olfactive, les mouvements et turbulences de l'air dans mon nez excitent de manière variable mes cellules réceptrices, mais je peux néanmoins sentir une même odeur. Cette possibilité pose évidemment problème. C'est ce qu'on appelle le " problème de l'invariance perceptuelle ".
Dans son ouvrage How Brains make up their mind (2000), Walter J. Freeman explore les bases neuronales de la perception sous l'angle du problème de l'invariance perceptuelle. Le texte qui suit retrace les grandes lignes des réflexions de Freeman sur la perception, et en particulier sur son objet d'étude particulier, la perception olfactive. Cela ne l'empêche toutefois pas de donner une portée générale à ses conclusions, les différentes modalités perceptuelles partageant, selon Freeman, le même type de mécanisme neuronal.
À
la recherche du pattern neuronal invariant correspondant à une odeur
Pour résoudre en terme neuronal le problème de l'invariance olfactive, il faut trouver dans les zones neuronales impliquées dans la perception olfactive une activité neuronale invariante pour un type de stimulus donné. Cette invariance neuronale ne se trouve évidemment pas chez les cellules réceptrices. En effet, la variabilité neuronale en réponse à la variabilité des différents modes de présentation des stimuli prend forme au sein même des cellules réceptrices. Si l'invariance n'est pas au niveau des cellules réceptrices, doit-on penser qu'elle se trouve au niveau de l'activité neuronale du bulbe olfactif, le second relais de l'olfaction?
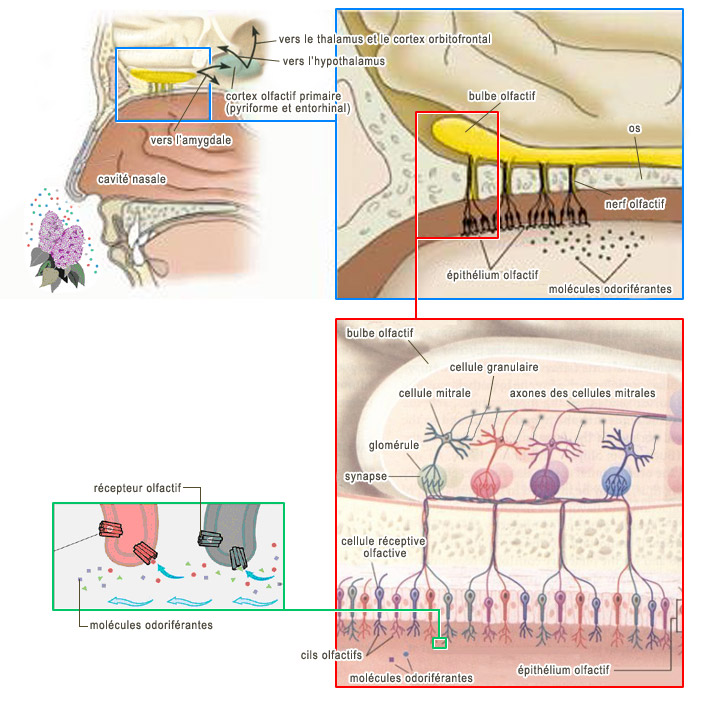
À première vue, il semble que la variabilité des patterns propres aux cellules réceptrices devrait se retrouver au sein des patterns d'activité du bulbe olfactif puisque les cellules réceptrices font des connexions directes aux neurones du bulbe olfactif. Et c'est effectivement ce que l'on observe, c'est-à-dire une grande variabilité dans l'activité neuronale du bulbe olfactif même si, contrairement aux cellules réceptrices, les neurones du bulbe olfactif interagissent entre eux.
Cependant, Freeman a remarqué que cette variabilité au sein du bulbe olfactif ne valait que si ses neurones étaient considérés individuellement, c'est-à-dire à l'échelle microscopique. Mais on peut discerner dans le cerveau deux autres échelles de grandeur : le niveau mésoscopique et le niveau macroscopique. Le niveau mésoscopique s'étend sur des surfaces du cerveau ayant quelques centimètres carrés; le niveau macroscopique, sur des aires complètes du cerveau. Au niveau mésoscopique, on tient compte de l'activité moyenne d'un très grand nombre de neurones individuels (microscopiques); au niveau macroscopique, de l'activité moyenne de l'activité d'ordre mésoscopique.
L'activité microscopique, avons-nous dit ci-dessus, ne contient pas de pattern d'activité invariant pour une odeur donnée. Selon Freeman, c'est au niveau mésoscopique que l'on trouve ce pattern d'activité invariant, ce que l'on voudrait tenter d'exposer ici. Mais avant de définir ce pattern d'activité invariant, il serait souhaitable de comprendre pourquoi, en passant de l'échelle microscopique à l'échelle mésoscopique, on délaisse le comportement microscopique (le comportement des individus) au profit du comportement moyen des individus.
Lorsqu'on passe d'une échelle de grandeur à une autre, on s'intéresse à l'activité moyenne des individus car, à l'échelle supérieure, seule l'activité moyenne possède une force causale. Afin de s'en persuader, l'exemple de la température est parfait. La température est une propriété macroscopique. Elle correspond à la vitesse moyenne des molécules d'un objet, et il ne fait aucun sens de considérer cette propriété relativement à l'activité microscopique (c'est-à-dire le mouvement individuel des molécules de cet objet), car nous ne sommes pas sensibles aux vitesses individuelles, mais seulement à la vitesse moyenne des molécules.
C'est une distinction du même ordre qui existe en électrophysiologie entre les enregistrements unicellulaires et extracellulaires. Dans le premier cas, on utilise une microélectrode pour pénétrer à l'intérieur d'un seul neurone. On peut ainsi enregistrer les potentiels d'action qu'émet ce neurone particulier. Dans le cas d'un enregistrement extracellulaire, l'électrode demeure cette fois dans le milieu extracellulaire et capte l'activité de plusieurs neurones avoisinants. Le tracé recueilli montre donc l'activité moyenne pour une région cérébrale donnée, sans que l'on sache de quel neurone précis provient chaque potentiel d'action de l'enregistrement.
Maintenant que l'on sait que l'échelle mésoscopique concerne l'activité moyenne des neurones individuels, revenons au problème qui nous intéressait initialement, c'est-à-dire décrire le pattern neuronal invariant (d'ordre mésoscopique) ou, autrement dit, le pattern neuronal qui reste le même malgré le fait que l'excitation venant des cellules réceptrices est variable.
Un pattern neuronal invariant apparaît dans le bulbe olfactif lorsque l'animal inhale une odeur particulière. Plus précisément, lorsqu'il y a inhalation de cette odeur, le bulbe olfactif dans son entier oscille à la même fréquence. Deux choses doivent être expliquées ici. D'une part, qu'est-ce qui fait que le bulbe olfactif oscille? D'autre part, qu'est-ce qui fait qu'il oscille partout à la même fréquence. Il oscille partout à la même fréquence parce que les neurones du bulbe possèdent un degré très élevé d'interconnexions.
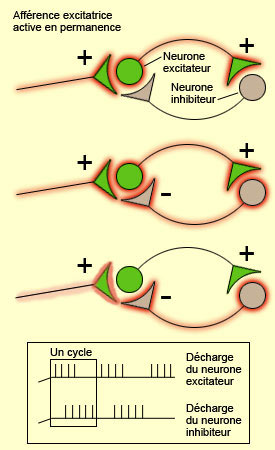
L'existence d'oscillations dans le bulbe olfactif est plus difficile à expliquer. Freeman soutient que l'oscillation provient d'un feedback négatif au sein d'une population mixte, c'est-à-dire une population formée de neurones excitateurs et inhibiteurs. Pour avoir une idée de la façon dont une population mixte qui interagit par feedback négatif peut produire une activité oscillatoire, nous reproduisons ci-dessous un encadré tiré du thème sur le sommeil, au niveau cellulaire avancé de notre site.
Différents types de connectivité neuronale favorisent l'émergence de bouffées rythmiques de potentiels d'action au sein d'un réseau de neurones. Ainsi, l'un des plus simples que l'on peut identifier est la connexion réciproque entre un neurone excitateur et un neurone inhibiteur, lesquels sont activés par un troisième neurone (dont le pattern d'activation peut être, lui, régulier et sans bouffées rythmiques). Aussi longtemps que dure cette activation constante du neurone excitateur en provenance de l'extérieur, l'activité de ce neurone excitateur va être périodiquement interrompue parce que celui-ci active le neurone inhibiteur qui l'inhibe en retour. L'arrêt temporaire de l'activité du neurone excitateur fait immédiatement cesser celle du neurone inhibiteur, ce qui rend à nouveau le neurone excitateur réceptif à l'activation constante de l'afférence extérieure. D'où l'activation du neurone excitateur qui reprend, entraînant rapidement celle du neurone inhibiteur, et la reprise du cycle oscillatoire.
|
|  |
L'oscillation qui couvre tout le bulbe ne possède pas partout la même amplitude. À certains endroits du bulbe, l'oscillation est plus forte; à d'autres, moins forte L'amplitude d'une population de neurones correspond au degré d'activation de la population, et le degré d'activation d'une population peut se mesurer par la fréquence des influx nerveux de la population. En raison de ces modulations d'amplitude, le bulbe forme un pattern de modulation d'amplitude ou pattern AM (pour " Amplitude Modulation ", en anglais). Ce sont ces patterns AM qui sont invariants face à la variabilité des stimulations provenant d'une même odeur. Pour chaque odeur excitant de manière variable les cellules réceptrices, on peut faire correspondre un seul pattern AM. Autrement dit, le pattern AM est en quelque sorte la " signature " neuronale de l'odeur. Le problème de l'invariance perceptuelle est donc apparemment résolu.
Une modélisation dynamique
de la perception olfactive
Le processus par lequel des inputs différents donnent lieu à un même pattern AM (c'est-à-dire l'essence du " problème de l'invariance perceptuelle ") est qualifié par Freeman de processus de généralisation. Si l'on comprend par " acte de généralisation " un acte où des entités différentes sont considérées comme semblables, on peut effectivement parler ici de généralisation, puisque des inputs, en soi différents les uns des autres, sont rapportés à la même chose, à un même pattern AM.
Freeman explique en termes dynamiques ce processus de généralisation. Autrement dit, il modélise le processus de généralisation en termes d'attracteurs et de bassins d'attraction. Un attracteur est un état (ou un cycle) qui se maintient dans le temps, et un bassin d'attraction est la région de l'espace d'états dont les états conduisent à l'attracteur.
Maintenant, comment Freeman modélise-t-il le processus de généralisation (ou, autrement dit, le processus par lequel des inputs variables mènent au même pattern AM) ? Freeman conçoit le pattern AM correspondant à une odeur comme un attracteur et les différents états du système (le bulbe olfactif) qui font immédiatement suite à l'inhalation comme des états du système qui se situent dans le bassin d'attraction de l'attracteur. Puisque ces états (qui correspondent aux patterns excitateurs variables) se situent dans le bassin d'attraction, ils conduisent inévitablement vers l'attracteur (le pattern AM). Puisque le bassin d'attraction et l'attracteur expliquent que des états variables mènent vers un même état (le pattern AM), ils expliquent du fait même le processus de généralisation.
Freeman modélise le bulbe olfactif comme un paysage d'attracteurs (" attractors landscape ", en anglais). Dans ce paysage d'attracteurs, on retrouve plusieurs bassins d'attraction ainsi que leur attracteur respectif. Chaque bassin et son attracteur définit ce que Freeman nomme une aile (" wing ", en anglais) du paysage d'attracteurs. Et chaque aile correspond à une odeur apprise. Parmi toutes les ailes qui composent le paysage d'attracteurs, il en existe deux qui sont particulières. La première est celle qui correspond à l'odeur d'arrière-plan, c'est-à-dire lorsque l'inhalation n'apporte pas d'odeur apprise. La deuxième aile correspond au moment qui sépare chaque inhalation, c'est-à-dire lorsque l'animal expire. Autrement dit, lorsque les cellules réceptrices ne sont pas excitées (entre les inhalations), le système tombe dans un bassin d'attraction particulier qui le mène vers un pattern AM particulier.
Maintenant que la possibilité d'une généralisation peut être comprise en termes dynamiques, on peut tenter de rendre compte de sa genèse. Comme Freeman explique la généralisation par l'existence d'un bassin d'attraction, cela revient à dire qu'il faudrait expliquer la genèse du bassin d'attraction correspondant à chaque odeur apprise.
La création d'un
bassin d'attraction grâce à l'apprentissage
La création d'un bassin d'attraction correspondant à une odeur apprise repose sur les processus neuronaux qui ont lieu lors de l'apprentissage. Avant de voir comment les processus neuronaux sous-jacents à l'apprentissage construisent un bassin d'attraction, il faut comprendre que Freeman s'intéresse en bout de ligne aux comportements- ce qui ne veut pas dire qu'il est behavioriste - et il détermine ce qui caractérise l'apprentissage dans cette perspective.
L'apprentissage auquel réfère Freeman est un apprentissage par renforcement. Pour apprendre à discerner une nouvelle odeur, il faut que l'animal ait associé cette odeur à un renforcement positif ou négatif. Le but de l'expérimentateur est alors le suivant : faire en sorte que l'animal (chez Freeman : un lapin) exécute une certaine action lorsqu'on lui présente une nouvelle odeur. Par exemple, l'action peut consister à lécher la substance odorante. Si l'animal parvient à lécher la substance odorante sur une base régulière, on pourra dire de l'animal qu'il a appris à discerner l'odeur.
Pour arriver à ce résultat, comme on l'a dit, Freeman doit renforcer l'action par un stimulus positif. Ainsi, si le lapin est assoiffé, Freeman présente rapidement au lapin un contenant rempli d'eau lorsque ce dernier a exécuté l'action appropriée (lécher la substance odorante). Maintenant, qu'est-ce qui nous fait dire que, à défaut de ce renforcement, l'animal ne parvient pas à discerner l'odeur? Il n'est pas impossible que ce renforcement soit seulement effectif pour l'action de l'animal; que l'animal sait déjà discerner la nouvelle odeur avant même de savoir exécuter l'action appropriée (lécher la substance odorante). Ce qui fait dire à Freeman que, à défaut de ce renforcement, le lapin ne discerne pas la nouvelle odeur, c'est que le pattern AM correspondant à l'odeur non renforcée est identique au pattern AM correspondant à l'inhalation de l'air d'arrière-plan, un air qui ne fait l'objet d'aucun discernement chez l'animal.
Revenons maintenant aux processus neuronaux sous-jacents à l'apprentissage et à la façon dont ils génèrent un bassin d'attraction. Comme on l'a dit, cette explication de la genèse du bassin d'attraction revient en fait à une explication de la genèse de la généralisation, puisque la possibilité de généraliser a été identifiée à la possibilité de poser un bassin d'attraction.
Pour qu'un bassin d'attraction soit créé, il faut que des états différents du système conduisent à un même état. Autrement dit, d'un point de vue neuronal, il faut que des patterns d'activité neuronale différents conduisent au même pattern d'activité neuronale. Pour que cela soit possible, il faut qu'un réseau fortement interconnecté soit établi ou, autrement dit, il faut qu'une assemblée neuronale soit créée. En effet, si une assemblée est construite, alors, compte tenu du degré élevé d'interconnexion entre les neurones de l'assemblée, l'activation de sous-ensembles différents mène à la même activation d'ensemble; ces activations différentes vont mener à l'activation de l'assemblée proprement dite, laquelle, lorsqu'on se place à un niveau mésoscopique, génère ou cause un pattern AM particulier dans tout le bulbe olfactif. Donc, en définitive, la création d'un bassin d'attraction équivaut à la création d'une assemblée neuronale. L'apprentissage devrait donc être en mesure de créer des assemblées neuronales. Il s'agit maintenant de montrer comment l'apprentissage crée de telles assemblées neuronales.
À chaque fois que l'animal inhale, un groupe de cellules réceptrices est activé. Ces cellules réceptrices vont ensuite activer des neurones du bulbe olfactif. Les neurones du bulbe olfactif, parce qu'ils sont interconnectés, peuvent renforcer leurs connexions synaptiques. C'est ce qui se produit lorsque deux neurones interconnectés font feu simultanément suite à une activation simultanée : les connexions synaptiques entre ces neurones sont renforcées (apprentissage hebbien).
En fait, le renforcement des connexions entre une paire de neurones ne se produit que lorsqu'il y a renforcement " comportemental " (par ex : lorsqu'on apporte de l'eau à l'animal assoiffé). C'est le relâchement d'un neuromodulateur par ce renforcement qui permet d'augmenter le poids des connexions entre les neurones. Ainsi, à chaque inhalation, des paires de neurones du bulbe font feu simultanément et le poids des connexions qui lient chacune de ces paires est augmenté (à condition, comme on l'a dit, qu'un renforcement " comportemental " suive immédiatement l'inhalation). Ce constat suggère que, à la suite d'une multitude d'inhalations, une quantité innombrable de neurones ont formé un réseau de neurones fortement interconnectés. Autrement dit, une assemblée neuronale a été créée.
Les processus neuronaux sous-jacents à l'apprentissage permettent donc effectivement de comprendre la genèse d'une assemblée neuronale. Et comme cette dernière définit un bassin d'attraction, on comprend comment l'apprentissage génère un bassin d'attraction ou, autrement dit, un processus de généralisation.
À la recherche d'une généralisation définitive
La généralisation effectuée dans le bulbe olfactif n'est pas définitive. Si la généralisation du bulbe était définitive, il faudrait que tous les inputs provenant d'une même source (une même odeur) donnent le même pattern AM dans le bulbe olfactif. Or, ce n'est pas le cas : dans le bulbe olfactif, le pattern AM correspondant à une odeur change avec le temps. Le temps n'est pas le seul facteur qui explique les changements du pattern AM. L'apprentissage d'une nouvelle odeur cause aussi ces changements. L'apprentissage d'une nouvelle odeur modifie les patterns AM existants car l'apprentissage d'un nouveau pattern AM implique que le paysage d'attracteurs " fasse de la place " à un nouveau bassin d'attraction, et ce changement de place signifie une modification des patterns AM existants. Il existe encore d'autres facteurs qui peuvent modifier les patterns AM du bulbe olfactif, mais nous ne les exposerons pas ici.
La question qui doit nous occuper est plutôt celle-ci : existe-t-il dans le cerveau un endroit où une généralisation définitive se produit; un endroit où il existe un pattern AM qui reste vraiment invariable? Et la réponse est oui. Ce pattern AM qui ne change pas à travers le temps (et à travers l'apprentissage de nouvelles odeurs) se trouve dans la région du cerveau vers laquelle les neurones du bulbe olfactif sont projetés, à savoir dans le cortex olfactif. En fait, la relation qu'un pattern AM du cortex olfactif entretient avec les patterns AM du bulbe est analogue à la relation qu'un pattern AM du bulbe entretient avec le pattern des cellules réceptrices. Le pattern AM du cortex olfactif est un attracteur pour les patterns AM du bulbe olfactif, tout comme le pattern AM du bulbe est un attracteur pour les patterns des cellules réceptrices. Autrement dit, on retrouve ici un processus de généralisation supérieur. Alors que le bulbe olfactif généralisait les patterns des cellules réceptrices, le cortex olfactif généralise les patterns AM du bulbe olfactif.
Merci à Martin Ratte pour sa contribution
à cette capsule.
|
|