Capsule outil: Le « chronomètre mental »
Notre
capacité à percevoir le temps qui passe a toujours intrigué
les scientifiques. Plus spécifiquement, ils se sont demandé si notre
habileté à sentir le temps qui s’écoule était
gouvernée par une horloge particulière dans le cerveau, ou bien
s’il s’agissait simplement d’un sous-produit de facultés
plus générales comme la mémoire ou l’attention.
Chose certaine, nous possédons tous un « chronomètre
mental », comme on l’appelle souvent, qui régit la perception
du temps dans une échelle qui s'étend de la seconde à la
minute. Ce chronomètre s’occupe donc d’une fenêtre temporelle
située entre l’horloge biologique de nos cycles circadiens qui suit
un cycle d’environ 24 heures, et toutes les oscillations nerveuses de l’ordre
des millisecondes associées à n'importe quelle activité de
notre cerveau.
Le chronomètre mental, pour sa part, vous indique à quelle vitesse vous devez courir pour attraper une balle de baseball. Ou quand claquer des doigts pour garder le rythme qui accompagne une pièce musicale. Ou encore combien de temps vous pouvez encore vous prélasser au lit après que le réveille-matin a sonné.
Il nous est également utile pour comprendre l'ordre temporel d’événements, par exemple lors d'une conversation. Pour comprendre le discours, le cerveau doit appréhender la durée des voyelles et des consonnes, rythmer la parole, organiser les pensées avec cohérence et répondre de façon opportune.
Autre exemple familier : quand un feu de circulation passe au jaune, on peut évaluer depuis combien de temps il est jaune. Puis comparer cette valeur avec le souvenir que l’on a de la durée d’un feu jaune. Finalement on décide si on a le temps de passer ou pas et on agit en conséquence. Finalement, on se rend compte qu’il serait difficile de trouver un seul processus comportemental complexe dans lequel notre chronomètre mental ne serait pas impliqué.
***
Le fonctionnement d’un tel chronomètre est forcément lié à plusieurs autres capacités cérébrales comme l’attention, qui permet de noter le début et la fin de l’intervalle de temps, et la mémoire, qui permet d’emmagasiner cette durée pour comparaisons futures.
Notre perception subjective du temps qui passe est d’ailleurs intimement liée au niveau d’attention que nous accordons à un intervalle de temps donné. Si cet article vous intrigue, le temps que vous passerez à le lire vous paraîtra court. S’il vous ennuie, au contraire, vous trouverez le temps long.
Cette durée subjective sera ensuite transférée dans la mémoire de travail où cette représentation pourra être maintenue et manipulée durant un certain temps, le temps par exemple de la comparer à une autre durée que vous venez d’évaluer, ou encore qui est stockée dans votre mémoire à long terme depuis longtemps.
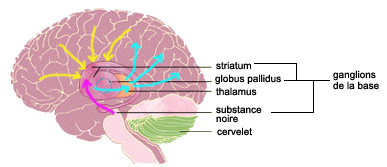
La localisation des circuits pouvant constituer un substrat neuronal à notre chronomètre mental est encore très débattue. Les ganglions de la base et le cervelet sont des candidats pressentis depuis longtemps puisque des lésions dans ces régions cérébrales perturbent des comportements nécessitant un calcul précis du temps. Mais comme ces comportements anormaux peuvent aussi être attribués à des dérèglements plus généraux du système moteur, la contribution de chacune de ces structures à notre capacité de percevoir les durées demeure ambiguë.
On a aussi proposé que l’activation dans les ganglions de la base pourrait survenir tôt dans le processus et être associée avec l’encodage des intervalles de temps, alors que l’activation du cervelet pourrait se faire dans un deuxième temps, suggérant une implication autre que la seule évaluation des durées.
D’autres études de lésions chez les animaux et les humains ont démontré que les lobes frontaux et pariétaux seraient aussi impliqués, mais d’une façon indirecte à travers leur rôle dans l’attention et la mémoire de travail.
Les sujets ayant subi des lésions à l’hémisphère droit ont aussi plus souvent des troubles d’évaluation des durées. Cette même tendance a été constatée dans des études d’imagerie cérébrale où une contribution supérieure de l’hémisphère droit dans les tâches de comparaison de durée a été remarquée.
***
La compréhension du mécanisme de notre chronomètre mental est très difficile parce qu’elle ne semble pas être localisée dans un secteur unique du cerveau, comme peut l'être l'horloge centrale du noyau suprachiasmatique, qui a des entrées claires situées dans la vision et qui déclenche l'émission cyclique des hormones et des comportements circadiens.
Mais au fur et à mesure que les données anatomo-fonctionnelles se sont accumulés, différents modèles ont été proposés pour tenter d’expliquer le mécanisme de notre chronomètre mental. Les circuits en boucle qui relient le cortex aux ganglions de la base ont ainsi inspiré un modèle de chronomètre où le calcul de la durée était déterminé grâce au temps que prenait l’influx nerveux pour faire un tour dans cette boucle. Il s’agissait donc d’une sorte de stimulateur biologique indépendant qui émettait un "tic-tac" à chaque fois qu’un cycle était complété dans la boucle.
Mais ce modèle, qui considère l’évaluation d’une durée comme quelque chose que le cerveau calcule indépendamment et ajoute ensuite à nos processus mentaux a été délaissé pour d’autres où le calcul de la durée est intrinsèquement lié à l’information de base en provenance des inputs sensoriels.
C’est ainsi que des chercheurs comme Warren H. Meck, de l’Université Duke en Caroline du Nord, ont développé une nouvelle approche de notre chronomètre mental basée sur la "détection de coïncidences" des oscillations de l'activité neurale.
L’utilisation de l’imagerie par résonance magnétique fonctionnelle (IRMf) a joué un grand rôle dans ces travaux. Elle a permis d’observer avec plus de précision les structures cérébrales impliquées dans l’évaluation des durées ainsi que la séquence temporelle de leur activation.
Ces techniques ont démontré que ce sont les ganglions de la base qui sont activés en premier lieu lors d’une tâche d'évaluation de durées. Et plus particulièrement la région du striatum qui possède une population de neurones richement interconnectés et recevant des signaux de nombreurses autres parties du cerveau. Les dendrites de ces neurones sont couvertes de 10 000 à 30 000 épines dendritiques, chacune d’elle recevant de l’information d’un neurone différent situé dans une autre région du cerveau. C’est d’ailleurs l’un des rares endroits dans le cerveau où l’on peut voir une telle convergence de milliers de neurones sur des neurones uniques comme ceux du striatum.
Le modèle proposé par Meck et d’autres chercheurs part du fait que les neurones du cortex ont des activités rythmiques très variées. Ainsi, beaucoup font feu de 10 à 40 fois par seconde spontanément sans recevoir aucune stimulation extérieure.
Chacun de ces neurones corticaux envoie donc des signaux aux dendrites des cellules du striatum qui intègre cette « musique neuronale » particulière. Arrive un événement particulier, par exemple le passage d’un feu de circulation au jaune, et les neurones corticaux font feu simultanément. Cela produit un « potentiel évoqué » caractéristique quelques 300 millisecondes plus tard. Ce potentiel évoqué agit un peu comme un coup de fusil de départ après lequel les neurones corticaux reprennent leur activité oscillatoire désordonnée.
Mais parce que les neurones corticaux ont tous été mis en phase par l’événement d’intérêt, la reprise de leur fréquence oscillatoire naturelle provoque chez les neurones du striatum vers lesquels ils convergent un pattern typique et reproductible dont la forme varie avec le temps qui s’écoule. C’est donc ce pattern singulier du temps qui passe après un événement particulier que les neurones du striatum utiliseraient pour mesurer le temps écoulé.
Retenir la durée d’un événement, comme par exemple l’intervalle de temps où un feu de circulation est au jaune, nécessiterait ensuite la contribution d’une autre structure cérébrale, la substance noire, souvent associée aux ganglions de la base. Celle-ci enverrait une décharge du neurotransmetteur dopamine suite à l’événement qui détermine la fin de l’intervalle (le passage du feu au rouge dans notre exemple).
Cet afflux soudain de dopamine permettrait aux neurones du striatum de retenir le pattern d’oscillations reçu à cet instant précis. Un peu comme une photo qui leur permettrait par la suite d’identifier un pattern semblable, et donc une durée équivalente. Car c’est bien ce que postule cette hypothèse, que chaque instant après le début de l’intervalle a une signature qui lui est propre.
Quand un neurone du striatum connaît ainsi la signature oscillatoire associée à la durée d’un événement particulier, quelque chose de différent se produira la prochaine fois que cet événement surviendra : une décharge de dopamine en provenance de la substance noire au début, et non à la fin, du potentiel évoqué des neurones corticaux associé au commencement de l’événement. Cet afflux de dopamine indiquerait aux neurones du striatum de commencer à analyser les patterns oscillatoires qui leur parviennent jusqu’à ce qu’il reconnaisse le pattern correspondant à la fin de la durée de l’événement.
À ce moment précis, un signal serait transmis du striatum au thalamus, qui communique pour sa part avec le cortex et les fonctions dites supérieures (mémoire, prise de décision, etc.). Le signal revient donc ainsi au cortex après son passage dans le striatum, mais cette fois-ci le cerveau sait combien de temps il lui reste avant la fin de l’événement. Dans le cas du feu de circulation, c’est alors que la personne peut décider de continuer ou d’appliquer les freins.
Le rôle de la dopamine dans ce modèle de chronomètre mental est appuyé par des expériences faites avec des gens qui produisent moins de dopamine comme les personnes atteintes de la maladie de Parkinson. Celles-ci sous-estiment en effet invariablement la durée des intervalles qu’on leur demande d’évaluer. Leur performance se corrige toutefois partiellement quand ces personnes sont traitées avec des médicaments qui augmentent le taux de dopamine dans leur cerveau.
À partir de la vingtaine, les taux de dopamine ont aussi tendance à diminuer avec l’âge. Cela pourrait contribuer au sentiment que le temps passe de plus en plus vite en vieillissant.
Le phénomène inverse semble aussi s’observer : des substances comme la cocaïne augmentent la disponibilité de la dopamine et provoque une accélération du chronomètre mental, de sorte que le temps nous apparaît plus long. L’adrénaline et d’autres hormones de stress accélèrent aussi le chronomètre mental ce qui expliquerait pourquoi quelques secondes peuvent nous apparaître interminables lors d’expériences désagréables.
Enfin, des états de concentration intense ou de grande émotion pourrait inonder ce système ou tout simplement le contourner, créant des moments où le temps semble suspendu ou ne pas exister du tout.
Comme les musiciens ou les athlètes le savent, ce chronomètre mental peut être entraîné pour l’amener à avoir une plus grande précision. Car sa précision chez le commun des mortels semble s’étendre de 5 à 60 % de la véritable durée. Et la marge d’erreur s’accroît avec la durée de l’intervalle. D’où l’aide précieuse que nous fournit l’objet que nous portons à notre poignet pour ne pas manquer nos rendez-vous…
Inspiré de : Times of Our Lives, By Karen Wright, Scientific American, September 2002.
|
|