|
|




L’hippocampe
entier révèle ses secrets
Beautés
de la nature… à tous les niveaux !
L’exercice
régulier : un remède contre l’anxiété
Dans les années 1970, une découverte
étonnante fit la démonstration que l'hippocampe du rat contient
une véritable carte spatiale de l'environnement où il évolue.
Certains neurones pyramidaux de la région CA1 vont en effet entrer en activité
uniquement lorsque le rat va se trouver dans une région précise
de son environnement. Ces cellules, dites "cellules
de lieu", sont au nombre de 1,000,000 dans l'aire CA1 du rat, de sorte que
si chacun se voit assigné un endroit précis dans l'espace, ils peuvent
former une carte cognitive très précise capable d'informer l'animal
à tout moment de l'endroit où il se trouve. De plus, l'exploration
d'un nouvel environnement va induire la formation d'une nouvelle carte cognitive
qui peut être très stable (semaines, mois). Une
des fonctions possibles de ces représentations spatiales dans l’hippocampe
serait, selon O’Keefe et Nadel qui les ont mis en évidence, de créer
un contexte sur lequel peuvent se greffer les souvenirs. Un événement
mémorisé peut alors devenir autobiographique, c’est-à-dire
situé dans le temps et dans l’espace, ce qui expliquerait le rôle
fondamental de l’hippocampe dans la mémoire
épisodique chez l’humain. | | |
| LA PLASTICITÉ DES RÉSEAUX DE NEURONES |
|
Aucun neurone isolé
ne contient en lui-même l'information nécessaire à la restitution
d'un souvenir. La trace mnésique est plutôt latente, ou encore virtuelle,
dans la mesure où son existence ne peut être mise en évidence
que lorsque qu'un réseau de plusieurs neurones interconnectés est
activé. Plusieurs souvenirs peuvent être
encodés à l'intérieur du même réseau de neurone
par différents patterns de connexions synaptiques. De même, un souvenir
peut aussi faire appel à l'activation simultanée de plusieurs assemblées
de neurones réparties dans différentes aires du cerveau. L'association
des ces groupes de neurones corticaux répartis dans différentes
aires cérébrales est rendu possible par certains réseaux
de neurones pré-câblés pour accomplir cette tâche. Parmi
eux, les circuits de la formation hippocampique, impliqués dans la l'établissement
de la mémoire
explicite à long terme, sont certainement les mieux connus de tous. L'information
en provenance des cortex associatifs visuels, auditifs ou somatiques parvient
d'abord à la région parahippocampique du cortex, puis passe au cortex
enthorinal et finalement à l'hippocampe
comme telle. À travers celui-ci, l'information va passer successivement
à travers trois régions distinctes. En
effet, l'hippocampe proprement dit est fait des régions pourvues de neurones
pyramidaux étroitement groupées, principalement les aires CA1, CA2
et CA3 (CA pour Corne d'Ammon, à cause de sa ressemblance avec la forme
des cornes de bélier du dieu Ammon de la mythologie). C'est ce que l'on
appelle le circuit ou la boucle trisynaptique de l'hippocampe.
| L'entrée de l'information
dans cette boucle unidirectionnelle se fait par les axones du cortex entorhinal
nommés fibres perforantes (ou voie perforante, car elle passe au travers
du subiculum et de l'espace qui le sépare du gyrus dentelé). Ces
axones font la première connexion de la boucle avec les neurones
du gyrus dentelé. Des cellules granuleuses du gyrus dentelé
partent les fibres moussues qui font la deuxième connexion de la
boucle avec les dendrites des cellules pyramidales de l'aire CA3. Les axones
des cellules de l'aire CA3 se ramifient en deux : une partie forme les fibres
commissurales se dirigeant vers l'hippocampe controlatéral via le corps
calleux et l'autre partie forme les collatérales de Schaffer qui font la
troisième connexion de la boucle avec les cellules de l'aire CA1. | 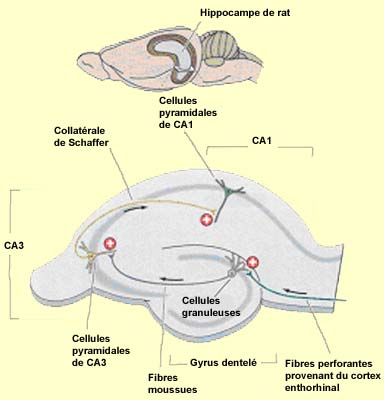 |
C'est dans ces synapses que la mémoire
spatiale associée à l'hippocampe semble être encodée
(voir encadré). Cette région démontre aussi une forte propension
à la potentialisation à long terme (PLT), bien que l'on observe
ce phénomène à plusieurs autres endroits dans l'hippocampe
ainsi que dans le cortex. Finalement, les axones des
cellules de CA1 se dirigent vers les neurones du subiculum et les neurones du
cortex entorhinal. La partie réceptrice de la formation hippocampique est
donc constituée par le gyrus denté et sa partie émettrice
par le subiculum. Les axones des grands neurones pyramidaux du subiculum se dirigent
ensuite vers les noyaux sous-corticaux par la fimbria, mince lame de substance
blanche au bord interne de l'hippocampe. Finalement, l'information retournera
vers les aires corticales sensorielles d'où elle était partie avant
d'être traitée par l'hippocampe.
|
|


